the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 20 May 2026
| 20 May 2026
Rock fracture characteristics regulate water storage and seasonal tree water uptake in karst
Xiuqiang Liu
Xi Chen
Zhicai Zhang
Weihan Liu
Tao Peng
Jeffrey J. McDonnell
The processes of tree water uptake in karst environments are poorly understood. One of the main challenges to improved understanding is the complex interaction between soil water and bedrock water, especially in systems characterized by structurally heterogeneous rock fractures. While some studies have highlighted the potential importance of fractured bedrock as a water source for plants, few have quantitatively assessed how fracture characteristics regulate water storage, residence times, and plant water uptake across seasons. Here, we combine stable isotope tracing, a Bayesian mixing model (MixSIAR), and hydrometric monitoring to quantify the contributions and mean residence times (MRT) of soil and rock water accessed by trees as a function of fracture properties. We use a four-compartment sampling framework that distinguishes between soil water (mobile and bulk) and rock water (fracture and infilled fracture). Our results show that fracture characteristics exert a primary control on seasonal tree water uptake patterns. During the peak growing season, mobile soil water (mean MRT =88 d) dominates uptake (mean contribution 41 %), whereas in late growing season, trees increasingly rely on bulk soil water (mean MRT =95 d, mean contribution 55 %). During the transition from the dry to the wet season, reliance on rock water increased in fracture-rich areas. In the reactivation stage, trees exhibited a mean rock-water contribution of 69 % (mean MRT =117 d), and in the subsequent early growing season, large trees derived up to 85 % of their water from rock, primarily from soil-filled fractures with apertures >10 mm that act as seasonal storage reservoirs with prolonged residence times (MRT =84–303 d). Trees preferentially access short-MRT sources under wet conditions and shift to longer-MRT pools during dry periods, demonstrating that seasonal water-use strategies are strongly regulated by fracture-controlled storage and connectivity. This study demonstrates that fracture characteristics play a central role in regulating rock water storage and tree water uptake in karst systems, providing new insights into vegetation resilience in structurally complex landscapes and implications for water resource management under changing climatic conditions.
- Article
(16664 KB) - Full-text XML
- BibTeX
- EndNote




Water transport and retention are critical for understanding the hydrodynamic behavior of the soil-plant-atmosphere continuum, in which the transfer of water through the plant and soil is a central process (Deng et al., 2017; Philip, 1966). Climate and critical zone regimes influence how plants access and utilize water resources by modulating precipitation inputs, root zone water transit, and residence times (Liu et al., 2024d; Luo et al., 2023; Fan and Miguez-Macho, 2024). Consequently, the temporal dynamics of water transport within trees directly influence how forests store and release water, thereby altering the timing and magnitude of water fluxes such as transpiration and runoff at the ecosystem scale (Bond et al., 2008).
Stable isotopes of hydrogen and oxygen (2H and 18O) have been powerful tracers for quantifying water transport (Putman and Bowen, 2019). When combined with isotope-based models, such as linear mixing models (IsoSource, SLM), Bayesian mixing models (SIAR, MixSIR, and MixSIAR), and continuous distribution models (e.g., CrisPy), the relative contributions of each water source to plant water use can be quantified (Wang et al., 2019a; Fu et al., 2024; Putman and Bowen, 2019). However, beyond mere source attribution, quantifying the temporal dynamics of water movement is essential for characterizing subsurface storage. Metrics such as Mean Residence Time (MRT) and Mean Transit Time (MTT) have been widely employed to estimate catchment water storage durations and the temporal lag between precipitation input and plant uptake (McGuire et al., 2002; Asadollahi et al., 2020; Liu et al., 2024d). While MRT reflects the storage capacity and mixing volume of a reservoir, and MTT characterizes the transport velocity, their application in complex karst environments remains challenging. Unlike homogeneous soils where these methods are effective (Liu et al., 2024d; Stewart and McDonnell, 1991), the high heterogeneity of karst fracture networks creates a dual-domain system characterized by rapid conduit flow and slow matrix storage. This complexity confounds standard residence time estimations (Hartmann et al., 2014; Zhang et al., 2021). Nevertheless, applying MRT and MTT analysis to specific karst water pools (e.g., fracture vs. matrix) offers a unique opportunity to unravel the temporal disparities that drive plant water availability.
Conventionally, isotope tracing tools have been applied to identify when and where soil water is accessed by plants (Midwood et al., 1998; Tang and Feng, 2001; McCole and Stern, 2007; Darrouzet-Nardi et al., 2006; Wang et al., 2010; Brooks et al., 2010). Recent advances have crystallized into the “ecohydrological separation hypothesis”, which posits that water contributing to plant transpiration is often distinct from the water that generates runoff or groundwater recharge (McDonnell, 2014; Brooks et al., 2010). This framework suggests the coexistence of at least two functionally distinct water reservoirs within the soil: a mobile, hydrologically connected fraction and a more strongly bound, plant-available fraction (Sprenger and Allen, 2020; Finkenbiner et al., 2022). However, empirical evidence for ecohydrological separation stems predominantly from soil-dominated ecosystems. In karst landscapes, the “soil” layer is often shallow and discontinuous, implying that separation mechanisms may fundamentally differ due to the underlying fractured bedrock. It remains unclear whether the ecohydrological separation hypothesis applies to the coupled soil–rock continuum, where the disparity between rapid fracture flow and water retained in soil-filled crevices could create a distinct form of separation critical for vegetation survival.
Following the seminal works of Salve et al. (2012) and Oshun et al. (2016) increasing attention has been directed toward a third, hidden water reservoir: rock moisture, defined as water stored in unsaturated weathered bedrock (Zhang and Zhang, 2021; Rempe and Dietrich, 2018). This water often exists at matric potentials above the turgor loss point of roots ( MPa) (Korboulewsky et al., 2020; Nardini et al., 2021; Zwieniecki and Newton, 1996; Schoeman et al., 1997; Hubbert et al., 2011). Under prolonged drought conditions, plants in specific environments are known to shift their water use strategy from shallow soil water to this deeper rock water (Ning et al., 2023; Jiménez-Rodríguez et al., 2022; Barbeta et al., 2015; Hahm et al., 2022; Nardini et al., 2024). Despite significant progress in delineating the geographical extent and importance of rock moisture (McCormick et al., 2021), the mechanisms of its extraction remain elusive. Since much of the weathered rock zone is fractured, few studies have quantified the specific contribution of rock fracture water to plant uptake.
It is well established that plant roots have the ability to access rock water in fractures (Schwinning, 2020). Studies suggest that the minimum fracture aperture required for root water uptake is approximately 0.1 mm (Zwieniecki and Newton, 1995; Schwinning, 2010). Once this threshold is exceeded, capillary forces diminish, while both hydraulic conductivity and volumetric flow increase significantly with fracture aperture (Liu et al., 2024c; Wang et al., 2015, 2022; Zimmerman and Bodvarsson, 1996). These hydraulically active microfractures can establish water connectivity across fracture networks (Wolfsberg, 1997), forming dynamic reservoirs (Jiménez-Rodríguez et al., 2022; Ning et al., 2023; Vrettas and Fung, 2017), that enhance the likelihood of plant access to rock moisture (Schwinning, 2020, 2010). Moreover, these fractures can act as sheltered conduits for root extension, buffering roots against environmental extremes such as thermal fluctuations and wind exposure, while their enclosed structure reduces evaporative losses, allowing water to be retained for longer periods (Pawlik et al., 2016; Zhang et al., 2016; Preisler et al., 2019; Hahm et al., 2020; Luo et al., 2024a). This protective effect allows roots to penetrate deep into the substrate and access water and nutrients that are inaccessible in shallow soils (Zhang et al., 2019a). Notably, larger fractures may become partially or fully filled with soil, further enhancing their capacity to retain water and provide nutrients to vegetation (Liu et al., 2024c; Peng et al., 2019; Estrada-Medina et al., 2013; Yang et al., 2016).
However, the mechanistic controls underlying plant water uptake from different fracture types remain poorly understood. Plants growing on different rock substrates may access varying amounts of available water, depending on the physical properties of the underlying fractures (Nardini et al., 2021, 2024; Querejeta et al., 2006; Zwieniecki and Newton, 1996). No study that we are aware of has yet systematically examined how fracture aperture, depth, and degree of soil infill influence both the residence time of stored water and the patterns of plant water uptake. We propose that these static physical characteristics of fractures are the primary drivers of dynamic seasonal shifts in plant water use. Rather than viewing seasonal variation solely as a climatic response, we argue that the structural heterogeneity of fractures (e.g., aperture and infill) dictates the temporal availability of water (residence time), thereby forcing plants to shift sources as different reservoirs fill or deplete at different rates. These structural attributes may determine whether a fracture functions as a fast conduit, a temporary buffer, or a long-term reservoir for plant-available water.
In this study, we conduct a systematic and mechanistic examination of the role of rock fractures on tree water use of water stored in bedrock. By incorporating within-fracture measurements and sampling, we target a representative karst landscape. In such karst environments, the epikarst serves as a pivotal hydrological interface between surface and subsurface systems, potentially regulating water storage, transmission, and plant growth (Wang et al., 2022). Epikarst thickness typically ranges from 0.5 to 30 m (Williams, 2008), and its highly developed pore and fracture networks not only facilitate the transformation of surface water into groundwater but also provide growth pathways for plant roots to access deep rock fractures. We focus on a site in southwest China's karst region, characterized by a subtropical monsoon climate. Ecologically, the site is dominated by mixed evergreen and deciduous broad-leaved secondary forests, which represent the typical vegetation recovery following karst rocky desertification in this region. Beyond its specific karst features, the Puding station serves as a natural laboratory for studying hydrological processes in shallow-soil, fractured-bedrock ecosystems globally. The site typifies complex Critical Zones where regenerating vegetation grows on thin, discontinuous soils overlie highly permeable bedrock (Zhang et al., 2019b; Jiang et al., 2020), ultimately increasing the frequency of drought events (Xu et al., 2023; Wang et al., 2019b). This bio-geological structure mirrors not only Mediterranean carbonate terrains but also other fractured rock environments where vegetation relies on rock moisture to survive seasonal drought (Wang et al., 2019b; Chen et al., 2011; Carrière et al., 2019; Liu et al., 2024a). Despite the hydrological challenges, the structure of the soil–rock system can modulate local water storage; in areas where the epikarst possesses stronger water-holding capacity, the combined soil-fracture system retains both water and nutrients, offering critical support to maintaining the stability and sustainability of these ecosystems (Wang et al., 2024b). However, the mechanistic extent to which this structural heterogeneity within the fractured bedrock dictates the partitioning of subsurface water resources and subsequently drives the water uptake patterns and survival of the overlying vegetation remains to be systematically unraveled.
We hypothesize that subsurface partitioning may extend beyond the soil matrix to include fractured bedrock, where water is stored in forms ranging from fast-draining open fissures to long-residence, soil-filled fractures. Although these subsurface compartments are hydraulically connected, they may differ substantially in renewal rates, retention times, and plant accessibility, effectively forming a structurally and functionally analogous extension of ecohydrological separation. This study aims to provide a scientific foundation for sustainable water resource management and vegetation restoration in karst environments. Specifically, this study will address three fundamental research questions:
- 1.
How do rock fractures (aperture, infill, and hydraulic characteristics) regulate water storage and plant water uptake?
- 2.
What controls the residence times of water across soil, rock, and plant compartments in karst systems?
- 3.
To what extent does ecohydrological separation occur within the coupled soil–rock system, and how is it modulated by seasonal rainfall?
To address these questions, we use a sampling approach where soil water is subdivided into mobile water (sampled via lysimeters) and bulk water (sampled via cryogenic extraction), while rock water comprises rock fracture water and infilled rock fracture water. This enabled us to distinguish among four types of water sources: mobile soil water, defined as water residing in connected soil pores that responds rapidly to rainfall events; bulk soil water, defined as more tightly bound water in micropores, characterized by slower renewal and longer retention; rock fracture water: defined as water stored in clean, unfilled fractures within weathered bedrock, often transient and dynamic and infilled rock fracture water, defined as water held in fractures partially or fully filled with soil, offering higher retention and potential nutrient supply.
2.1 The study area and sampling location
Our experimental site is located at the Puding Karst Ecosystem Research Station in Guizhou, southwest China (105° 42′–105° 43′ E, 26° 14′–26° 15′ N) (Fig. 1). The climate is dominated by subtropical humid monsoon. The mean annual temperature is 15.1 °C, and the mean annual precipitation is 1378 mm. Precipitation in this region occurs exclusively in the form of rainfall. Over 80 % of annual precipitation occurs from May to October as summer rainfall (Liu et al., 2022). The carbonate rocks consist mainly of limestone. Soils developed from black and yellow limestone are distributed irregularly across the study site (Li et al., 2023) with soil-filled “grykes” (i.e. cracks that form vertically in the limestone via chemical weathering, Parry, 1960) widely distributed (Fig. 1). The thickness of the root-zone soil ranges from 15 to 200 cm (mean: 66 cm), the bare rock coverage varies from 0.42 to 0.94, and the apertures of rock fractures range from 0.1 to 38 mm (mean: 2.4 mm), with over 70 % of fractures having apertures narrower than 1 mm. Fractures with apertures greater than 10 mm are often completely or partially filled with soil (Liu et al., 2024c, 2025). The vegetation at the site consists of secondary broadleaf forests that have regenerated after environmental disruption caused by human activities, such as deforestation for cultivation.
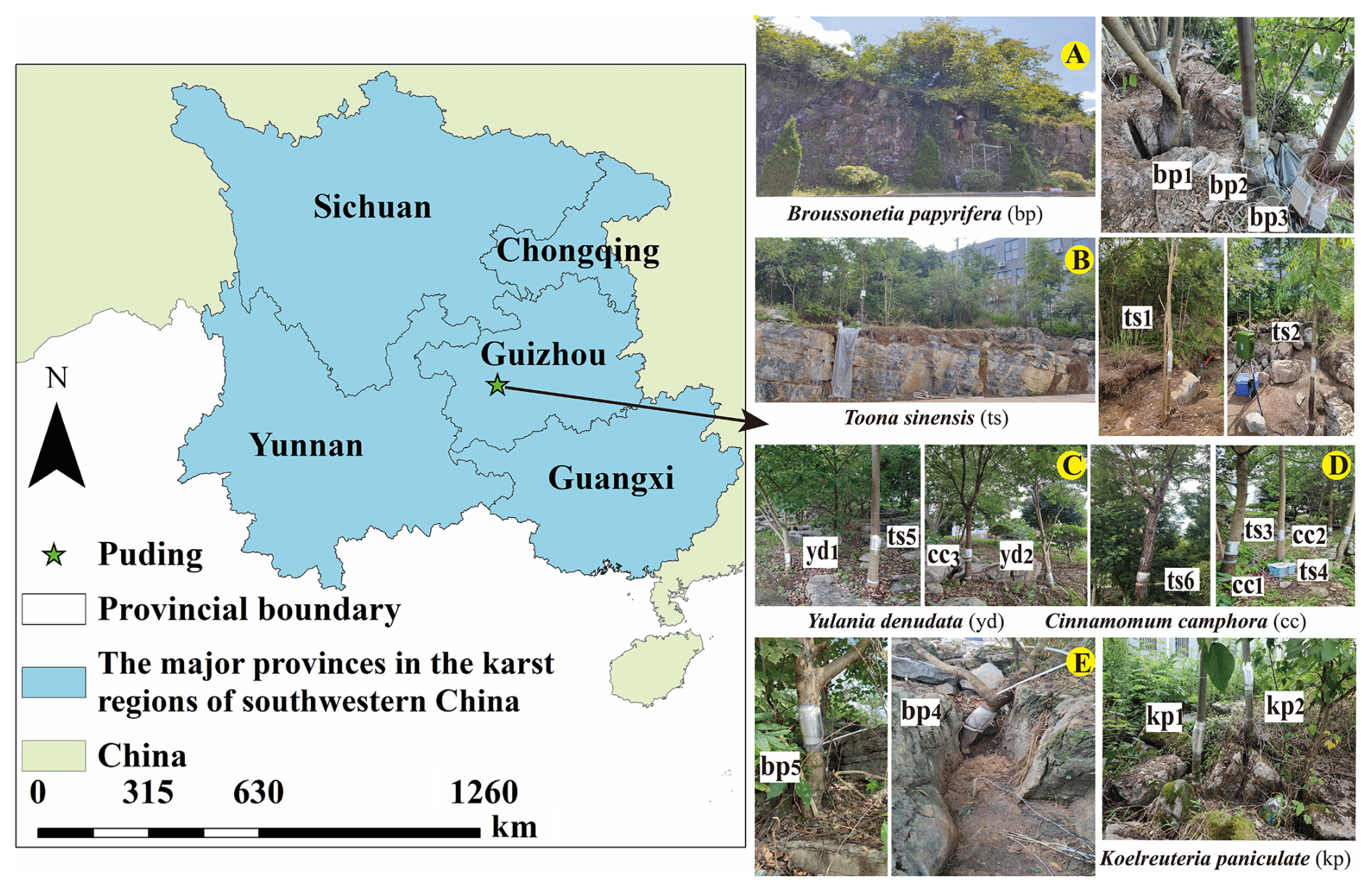
Figure 1The study area and five sampling sites.
We selected five typical sites (A–E in Fig. 1) to survey the soil–rock structure and tree stand structure (Liu et al., 2025). The five sites included 18 individual trees of 5 different species, including deciduous broadleaf trees (Broussonetia papyrifera (L.) L'Hér. ex Vent. (bp1–5), Koelreuteria paniculata Laxm. (kp1–2), Toona sinensis (A.Juss.) M.Roem. (ts1–6), and Yulania denudata (Desr.) D.L.Fu. (yd1–6)), and one evergreen broadleaf tree (Cinnamomum camphora (L.) J.Presl. (cc1–3)).
2.2 Field measurements
2.2.1 Collecting samples for isotope analysis
The method for collecting mobile soil water and bulk soil water samples at different depths involved two preparatory steps: (1) soil boreholes were manually drilled using a soil auger to predefined discrete depths at each study site; (2) at selected depths, soil material was removed and ceramic tensiometer tips connected to suction lysimeters were installed for the collection of mobile soil water. The boreholes were then backfilled with the original soil to ensure proper contact and minimize disturbance. Bulk soil samples were collected at all predefined depths and bulk soil water was subsequently extracted from the collected soil samples using laboratory extraction methods. The site-specific sampling depths, sampling frequency, and sampling periods for bulk soil water and mobile soil water are summarized in Table A1.
Following the conceptual framework of Sprenger et al. (2018), we distinguish between mobile soil water and bulk soil water based on their retention characteristics and accessibility under different matric suctions. Mobile soil water refers to the fraction of soil water that is weakly retained in the soil matrix and occupies larger, well-connected pores, making it accessible at relatively low matric suction and responsive to recent precipitation inputs. In this study, bulk soil water was sampled at least two days after rainfall, when gravitational water had largely drained from the soil profile. Under these conditions, the extracted bulk soil water was dominated by more strongly retained, matrix-bound water, with only a minor contribution from mobile water. To operationalize this conceptual distinction, soil water retention curves were constructed for each site and soil layer based on previously established soil hydraulic parameters (Liu et al., 2024c). Based on soil moisture distribution characteristics, site- and depth-specific upper suction thresholds associated with mobile soil water were identified for different soil types. The estimated maximum suction values corresponding to mobile soil water varied across sites and depths, reflecting differences in soil texture and hydraulic properties. Specifically, upper suction thresholds ranged from approximately 70–90 kPa at Site A (50–100 cm), 65–90 kPa at Site B (20–180 cm), 40–45 kPa at Sites C and D (20–50 cm), and 55–80 kPa at Site E. During field sampling, mobile soil water was collected using suction lysimeters operated at vacuum pressures below these site- and depth-specific upper suction thresholds to ensure that the extracted water predominantly represented mobile soil water rather than tightly bound soil water.
Rock fracture water and infilled rock fracture water were distinguished based on fracture aperture and the presence of infill material observed during drilling and coring. Fractures with apertures ranging from approximately 0.1 to 10 mm generally exhibited minimal soil or sediment infill and were therefore considered to predominantly store free fracture water. Water collected from these fractures is referred to as rock fracture water. In contrast, fractures with apertures larger than approximately 10 mm were commonly partially or fully filled with soil or fine sediments. Water collected from these zones primarily represents water stored within infilled fracture material and is referred to as infilled rock fracture water. For the collection of fracture water, 26 representative fracture locations were selected based on prior field investigations (Liu et al., 2024c). At each location, boreholes were drilled and rock cores were extracted using a portable core drilling tool to directly identify fracture geometry, aperture, and infill conditions. Ceramic tensiometer tips connected to suction samplers were positioned directly within the targeted fracture zones according to the fracture classification described above. To prevent mixing of water from fractures at different depths, PVA sponges were used to hydraulically isolate individual fracture layers within the boreholes. The ceramic tips installed in soil and fracture zones were connected to sampling bottles, and a vacuum pump was used to apply negative pressure for water extraction (Fig. A1). The site-specific sampling depths, fracture apertures, sampling frequency, and sampling periods for rock fracture water and infilled rock fracture water are summarized in Table A2.
At each study site, representative trees were selected for xylem water sampling. Tree physiological traits and root-zone soil–rock structural characteristics are summarized in Table A3. More detailed information on vegetation characteristics and root-zone soil–rock structure at the study sites is provided in Liu et al. (2025). For each sampling campaign, three branches were collected from different positions of each sampled tree to provide within-tree replication. Sampling was conducted from September 2022 to March 2024, with an average sampling frequency of approximately two to three times per month. The bark and phloem were removed immediately after sampling, retaining only the xylem tissue, and samples were promptly sealed in airtight containers prior to laboratory analysis.
Precipitation samples were collected during individual rainfall events using a standard precipitation collector installed at 1.5 m above ground level to minimize splash contamination and evaporation effects. The collector was placed in an open area, away from buildings and vegetation, to avoid external contamination. From September 2021 to March 2025, a total of 182 precipitation samples were collected.
All soil and xylem samples were stored frozen, while soil water, rock water, and rainwater samples were refrigerated for storage.
2.2.2 Moisture and sap flow monitoring
Meteorological variables were observed at the standard meteorological station of the Puding Karst Ecosystem Research Station. Automatically recorded variables included precipitation (P), air temperature at 2 m height (Ta), and soil moisture content at depths of 0–200 cm at each sampling site, monitored using capacitance-based frequency domain reflectometry (FDR) sensors (5TM, METER Group, Pullman, WA, USA) at 30 min intervals. Additional sensors at various depths were also installed at the soil–rock interface profiles at sites A and B. Detailed procedures for monitoring rock moisture content and sap flow in trees are described in Liu et al. (2025).
2.3 Laboratory analyses
Due to the long-term nature of the project and the development of international collaboration, sample processing and testing were divided into two analytical periods. Samples collected between September 2022 and March 2024 were processed and analyzed at the Ecohydrology and Water Resources Research Center and the Experimental Test Analysis Science and Technology Center, School of Earth System Science, Tianjin University. Samples collected from April 2024 to March 2025 were processed and analyzed at the University of Saskatchewan, Canada.
To collect xylem water from vegetation, the collected arboreal branches were processed using the LI-2100 fully automatic vacuum condensation extraction system (Beijing Ligajoint Scientific Co., Ltd., China) to extract liquid water (cryogenic vacuum distillation technique, CVD). Prior to extraction, the samples were weighed with a microbalance (accuracy of 0.1 mg), and the heating temperature for extraction was set to 150 °C with a duration of three hours. After extraction, the samples were weighed again to calculate the volume of water extracted, and then dried at 105 °C for 24 h. The dry samples were weighed again to calculate the total water loss, and the water extraction rate was calculated based on the weight loss (the proportion of water extracted to the total water loss after drying). The water extraction rate for all samples was generally above 99 %. Samples with less than 99 % water extraction rate were re-extracted until the standard was met.
Water isotopes δD and δ18O in rainfall, mobile soil water, and rock water samples were measured using the Picarro L2140-i water isotope analyzer. The measurement errors for δD were 0.1 ‰, and for δ18O were 0.015 ‰. As tree branch water contains volatile organic compounds (VOCs), all plant extracted waters were analyzed using the MAT 253 Plus gas stable isotope ratio mass spectrometer (IRMS) to determine δD and δ18O values. The errors for δD were 2 ‰, and for δ18O were 0.2 ‰.
Samples from April 2024 to March 2025 were processed and analyzed at the University of Saskatchewan in Canada.
Water was extracted from plant xylem and soil samples using the CVD method developed by Koeniger et al. (2011). After freezing the sample vials with liquid nitrogen, they were connected via a capillary tube to a collection vial, vacuumed to below 0.8 mbar, and heated at a predetermined temperature for a specified duration. In this closed system, vaporized water from the sample vial was condensed into the collection vial. The extracted water was then transferred to 2 mL screw-cap glass vials. Extracted samples were subsequently oven-dried at 105 °C for 24 h to evaluate extraction efficiency. Plant samples were extracted at 200 °C for 24 min, while soil samples were extracted at the same temperature for 30 min (Wang et al., 2024a). Higher extraction temperatures and efficient water recovery help to minimize isotope ratio errors in soil and xylem water samples (Younger et al., 2024).
Rainfall, mobile soil water, and rock water samples were analyzed using an Off-Axis Integrated Cavity Output Spectroscopy (OA-ICOS) system from Los Gatos Research, with post-processing performed via the LIMS for Lasers 2015 software. Due to the presence of organic compounds in plant and soil-extracted water samples, these were analyzed using IRMS. For hydrogen isotope analysis, the elemental analyzer-IRMS (EA-IRMS) method was employed, where water samples reacted with elemental chromium at high temperatures to produce hydrogen gas, following the method of Morrison et al. (2001). The resulting hydrogen gas was separated using a gas chromatographic column and then analyzed by IRMS. For oxygen isotope analysis, the CO2-H2O equilibration method described by Epstein and Mayeda (1953) was used, wherein water equilibrates with CO2 at a controlled temperature and the isotopic composition of the equilibrated CO2 is then measured by IRMS.
The analytical uncertainties (2σ) for LGR measurements of liquid samples were ±2 ‰ and ±0.8 ‰ for δD, δ18O, respectively. Water extracted from soil and plant samples was analyzed using IRMS. The analytical precision of IRMS was approximately ±1.0 ‰ for δD and ±0.2 ‰ for δ18O. All isotope values were reported in per mil (‰) relative to the Vienna Standard Mean Ocean Water-Standard Light Antarctic Precipitation (VSMOW-SLAP) scale.
Inter-laboratory consistency was further evaluated using overlapping precipitation isotope records, which showed no systematic offset between analytical periods.
2.4 Data analysis
2.4.1 Estimation of mean transit time and mean residence time
The mean residence time (MRT) represents an effective, isotope-based measure of the average time that water resides within a given compartment before being transferred to downstream compartments. Rather than describing a precise physical storage duration, MRT characterizes the seasonally integrated effects of storage, mixing, and transport processes. In karst environments, where flow pathways are highly heterogeneous, MRT is particularly useful for describing the dominant seasonal storage behavior of water within soil, fracture, and rock compartments. For example, soil water at 50 cm depth may retain infiltrated rainfall for a certain period before contributing to deeper flow or evapotranspiration. We estimated MRT using the amplitude damping approach (Małoszewski and Zuber, 1982; Reddy et al., 2006; Stewart and McDonnell, 1991; McGuire et al., 2002), which interprets the attenuation of seasonal isotope signal amplitudes as a response to mixing and storage along flow pathways. This approach assumes that seasonal isotope variability provides an integrated signal of water storage and turnover at the timescale of interest, which is well suited for investigating seasonal water storage dynamics in karst systems.The isotope sine curve analysis is used to estimate MRT by fitting a sinusoidal function to the isotope data of each compartment. We used the sine function fitting:
Where, a1 and a2 are the amplitude parameters, and offset represents the baseline value.
We applied a sine function transformation to the isotopic time series (δ18O or δD) as follows:
Amplitude (A) and phase (ϕ) were calculated as:
The MRT (in days) was calculated by:
Where An and Am are the amplitudes of the output and input isotope signals, respectively. The damping factor (f) was used to describe how the amplitude of the isotope signal is reduced as it moves through different compartments.
The isotope time series used for the estimation of MTT and MRT spans the entire period from September 2022 to March 2025 and therefore includes data analyzed in both laboratories. As shown by the precipitation isotope record (Fig. 2) and the isotopic dynamics of soil water, rock water, and xylem water (Figs. B1–B3), the isotope signals over this period exhibit smooth and continuous seasonal variations with clear sinusoidal characteristics. No discontinuities or step changes are observed at the transition between the two analytical phases, indicating good continuity and consistency between datasets obtained from the two laboratories. This temporal coherence results in stable sinusoidal fitting and low uncertainties in the estimation of MTT and MRT.
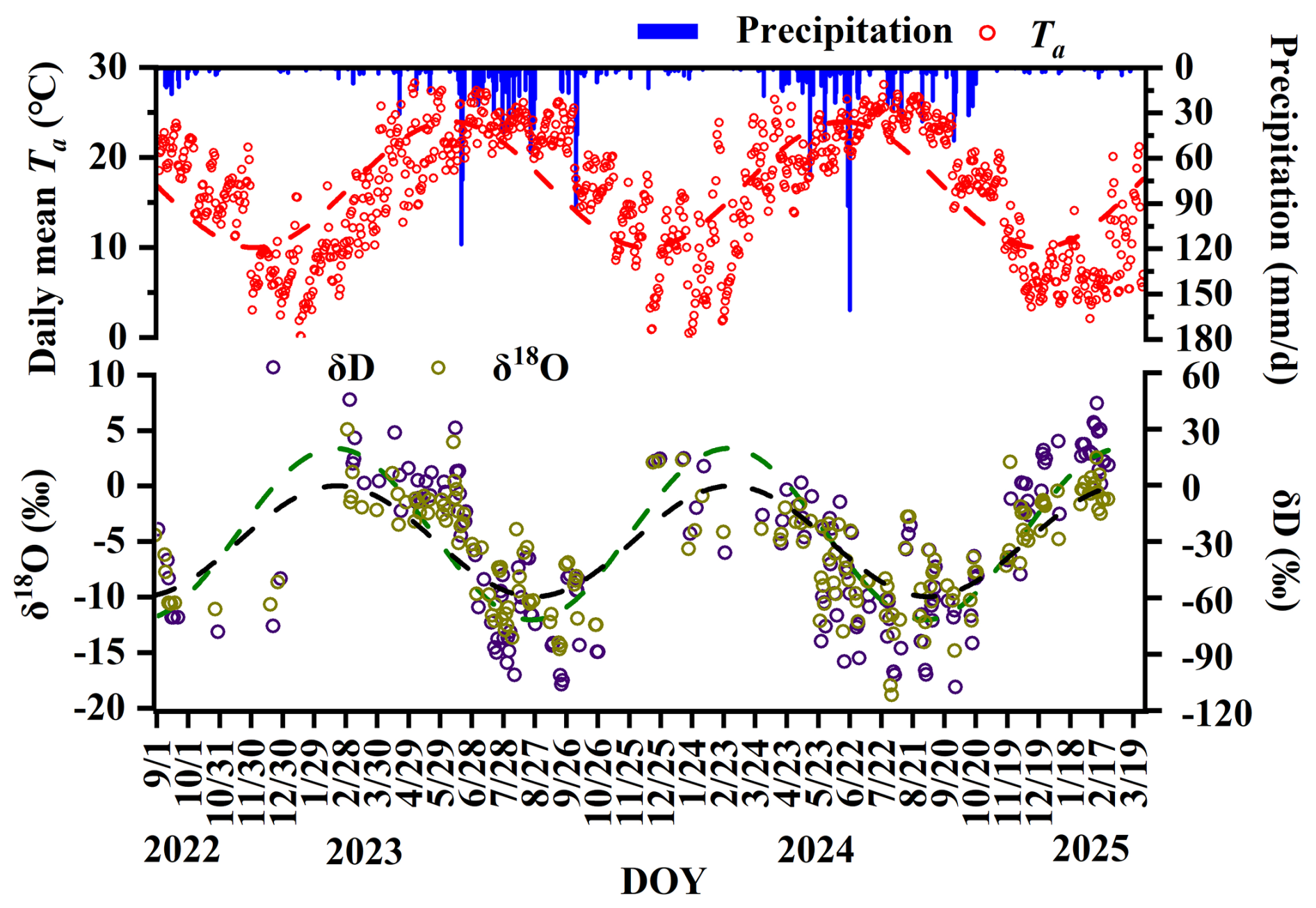
Figure 2Temporal variations in stable isotopes of precipitation (δD and δ18O), rainfall amount, and daily average temperature.
2.4.2 MixSIAR model
To analyze the partitioning of water sources among plants in our karst ecosystem, we employed the MixSIAR Bayesian Mixing Model (BMM). By integrating isotopic data, we gained a comprehensive understanding of the contributions of various water sources to plant water uptake. MixSIAR is a stable isotope analysis package in R that estimates water source contributions through a Bayesian approach, using Markov Chain Monte Carlo (MCMC) methods to generate posterior distributions of source proportions. The MCMC algorithm runs on multiple chains to ensure convergence and reliable posterior estimates (Moore and Semmens, 2008; Stock and Semmens, 2016; Parnell et al., 2013). One of the main advantages of MixSIAR over traditional mass balance methods is its reduced sensitivity to isotopic fractionation effects (Evaristo et al., 2017), and it incorporates uncertainties from the sources by introducing prior information, multiple continuous covariates, and error structures to improve prediction accuracy (Gai et al., 2023; Liu et al., 2024b). This robustness is particularly well-suited to karst ecosystems, where the isotopic composition of water sources can vary significantly due to complex interactions among soil, rock fractures, and vegetation (Gai et al., 2023).
The quantitative water source partitioning using the MixSIAR model is based exclusively on samples collected from April 2024 to March 2025. All samples used for source partitioning were processed and analyzed at the University of Saskatchewan, Canada. This analytical design minimizes potential inter-laboratory variability and ensures internal consistency among water sources, particularly for bulk soil water, which was extracted and analyzed only in Canada.
2.4.3 Calculation Method of Evaporation Index
The evaporation degree of different water bodies relative to local precipitation is characterized using the linear offset (Lc-excess) from the Local Meteoric Water Line (LMWL), as proposed by Landwehr and Coplen (2006):
where a and b are the slope and intercept of the LMWL, respectively.
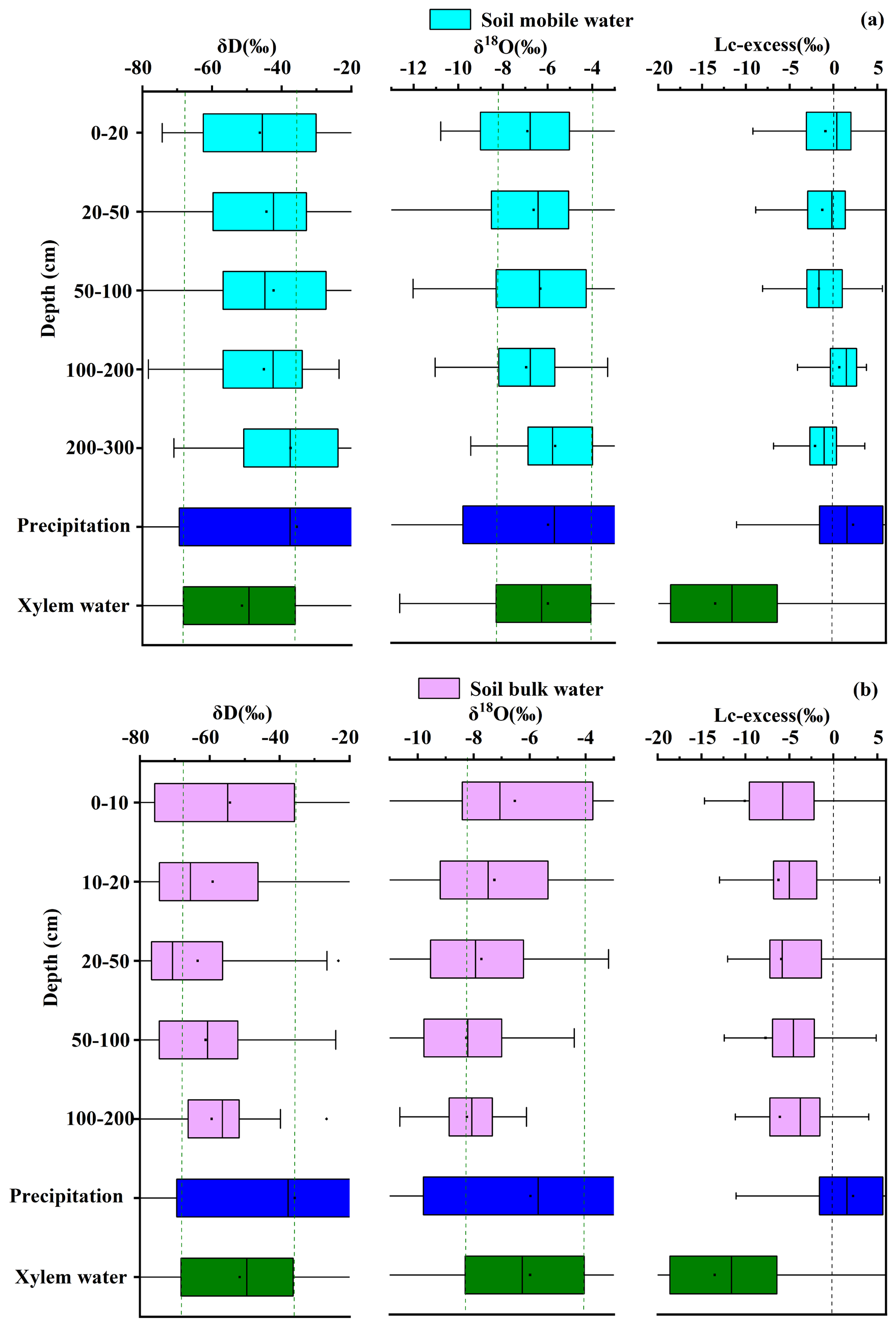
3.1 Temporal variation in isotope values and water conditions within water pools
The isotopic signatures of different water pools within the karst critical zone exhibited clear spatial and temporal variability, reflecting the contrast between dynamic precipitation inputs and more buffered subsurface reservoirs.
As shown in Fig. 2, the δ18O and δD values of precipitation displayed pronounced seasonal fluctuations that followed changes in precipitation amount, temperature, and moisture source. During summer and early autumn (June–September), abundant rainfall associated with maritime air masses resulted in relatively depleted isotopic values. After major rainfall events, isotope values decreased rapidly. This depletion is consistent with the combined influence of the amount effect, convective mixing, reduced sub-cloud evaporation, and shifts in moisture source regions. In contrast, precipitation during winter and spring (December–April) was mainly influenced by continental air masses and lower precipitation amounts, resulting in comparatively enriched δ18O and δD values.
The arithmetic mean values of precipitation (mean ‰, ‰) were used to represent the central tendency of the seasonal signal. The Local Meteoric Water Line (LMWL: ) closely overlapped with the Global Meteoric Water Line (Fig. 3), suggesting that precipitation isotopes in the study region are primarily controlled by moisture source rather than significant evaporative fractionation during atmospheric transport.
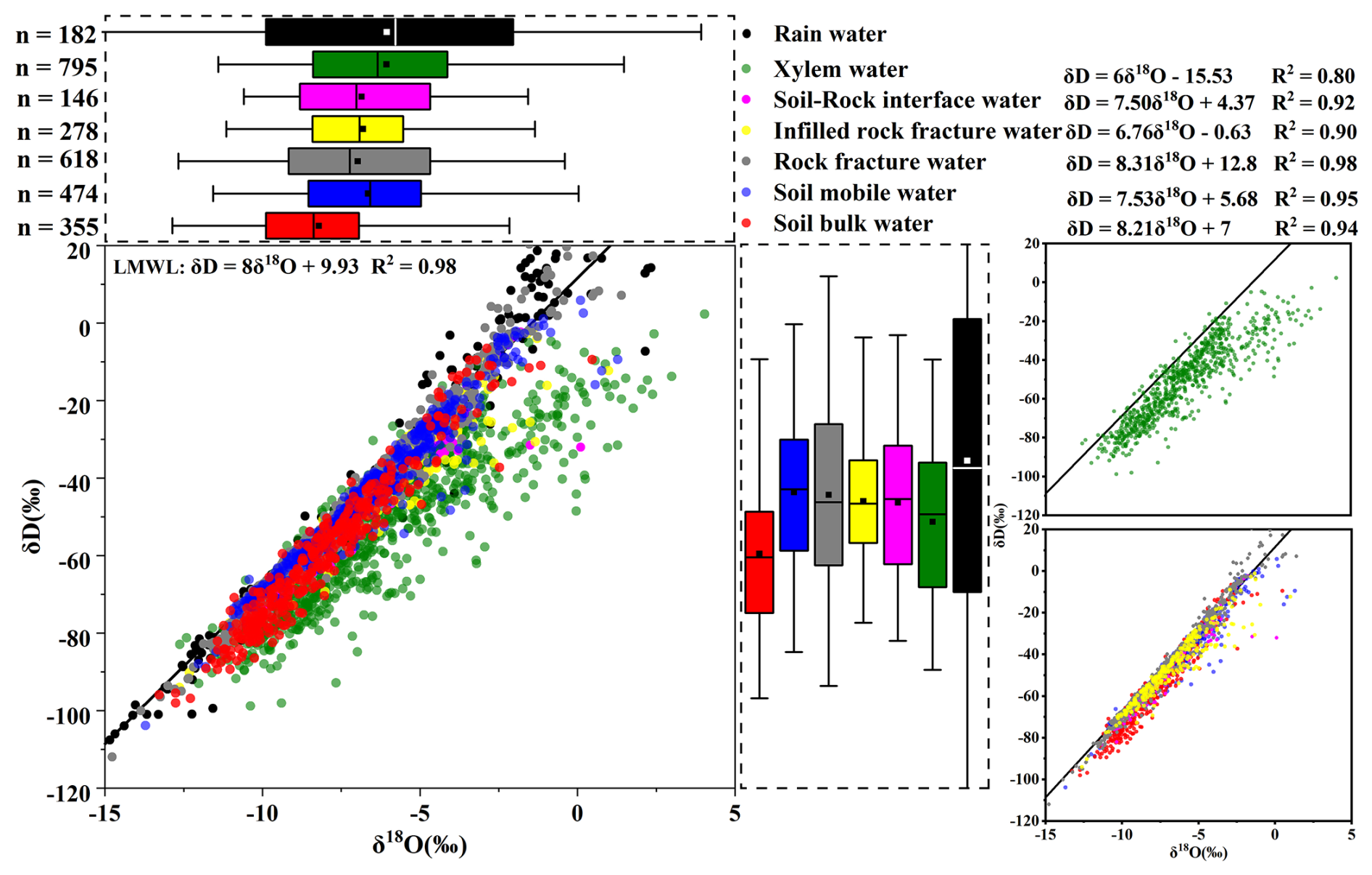
After infiltration, the isotopic compositions of subsurface water pools diverged from those of precipitation due to the structural heterogeneity of the soil–rock system. Mobile soil water (mean ‰, ‰) exhibited a relatively wide isotopic range and clear seasonal fluctuations that broadly followed the precipitation signal (Fig. B1a and b). These variations likely reflect the combined effects of depleted rainfall inputs during the wet season, evaporative enrichment during dry periods, and rapid mixing within the shallow soil profile. In contrast, bulk soil water showed more depleted mean isotopic values (mean ‰, ‰) and a more damped temporal signal. While shallow bulk soil water (0–30 cm) displayed noticeable fluctuations associated with evaporation and recent rainfall inputs, deeper soil water (>30 cm) remained relatively stable in both isotopic composition and moisture content (Fig. B1c), indicating slower water turnover and stronger mixing of older water.
Hydrological behavior at the soil–rock interface differed from that of the surrounding soil matrix. Soil-rock interface water (mean ‰, ‰) responded more rapidly to rainfall events and showed larger moisture fluctuations than adjacent soil water (Fig. B1d). This pattern indicates that the interface likely functions as a preferential flow zone rather than a slowly percolating matrix domain.
The isotopic variability of rock fracture water also depended on fracture characteristics. Rock fracture water (mean ‰) and infilled rock fracture water (mean ‰) exhibited different temporal stability depending on fracture aperture and the presence of soil infill (Fig. B2a and b). Shallow and narrow fractures (e.g., 45–60 cm at site B) showed relatively large fluctuations, suggesting rapid recharge from recent rainfall. In contrast, deeper fractures (e.g., 244–306 cm at site C) and large fractures filled with soil (e.g., 10–21 mm at site B and 38 mm at site C) exhibited more stable isotopic signals and weaker moisture fluctuations (Fig. B2c). These observations suggest that soil infill can reduce rapid infiltration and enhance water retention within fractures, allowing them to function as relatively stable subsurface water reservoirs.
Plant xylem water isotopes (overall mean ‰, ‰) reflected the integration of these different subsurface water pools. Xylem water samples plotted below the LMWL and aligned along a soil water line with a slope of approximately 6 (Fig. 3). This offset likely reflects isotope fractionation associated with soil evaporation and the mixing of different source waters within the soil–rock system. Seasonal variations in xylem isotopes broadly followed the availability of different water sources (Fig. B3a and b) but also exhibited clear spatial differences among sites. For example, trees at site B showed relatively enriched isotopic values (mean ‰, ‰), consistent with the isotopic composition of local water sources. In contrast, trees at sites A and C and D showed more depleted xylem isotopic signatures (mean ‰ and −6.17 ‰, respectively). These spatial differences suggest that vegetation water uptake is strongly influenced by local hydrological conditions, including soil water availability, root distribution, and access to different subsurface water pools.
3.2 Factors affecting MRT in different water pools
The residence times of soil water and soil–rock interface water were fundamentally controlled by permeability and depth, reflecting a clear divergence between stable storage and rapid preferential flow paths. Overall, bulk soil water exhibited significantly longer residence times (mean MRT =196 d) than mobile soil water (mean MRT =135 d) (Table C1). This divergence was strongly regulated by soil hydraulic conductivity (Kh). As shown in Fig. 4, the MRT of bulk soil water decreased exponentially with increasing Kh, stabilizing when Kh exceeded 0.75 cm h−1. In deep, low-permeability soils (e.g., >50 cm at site B, where Kh ranges only from 0.1 to 0.18 cm h−1, bulk water MRT extended up to 708 d (mean 513 d)), functioning as a slow-turnover storage pool. Conversely, the water at the soil–rock interface renewed rapidly (mean 118 d compared to 513 d for adjacent bulk soil; Table C2). These contrasting patterns demonstrate that while the soil matrix retains water long-term, deep mobile water is rapidly replenished via preferential flow along soil–rock interfaces.
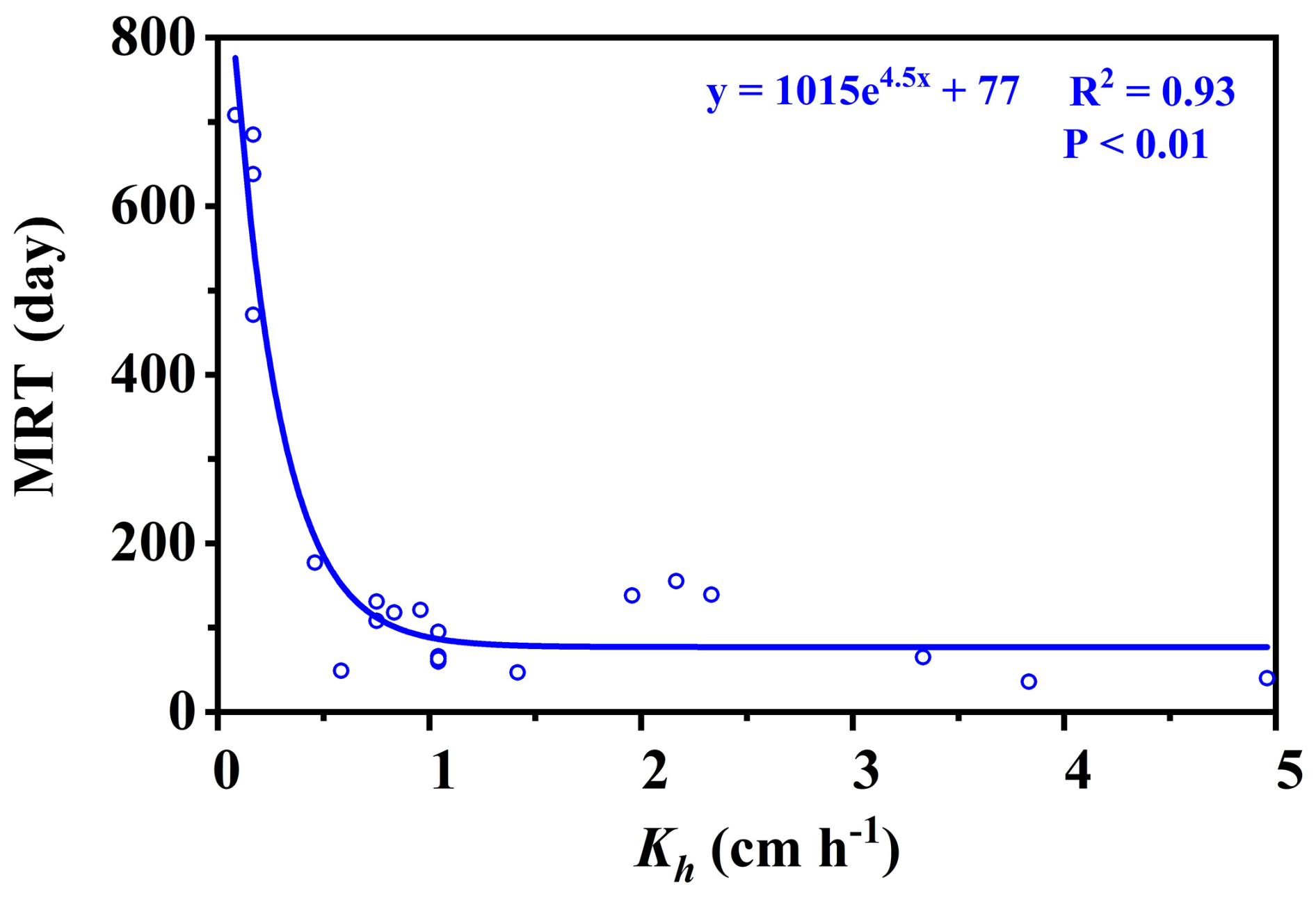
Crucially, the retention capacity of bedrock water was directly dictated by fracture architectural traits, specifically aperture size and the presence of soil infill (Fig. 5). While shallow, narrow fractures (aperture 0.47–2 mm, depth 45–80 cm) exhibited rapid renewal driven by precipitation (MRTs of 23–91 d), deeper and wider fractures showed pronounced storage capabilities (Table C3). A significant positive threshold relationship emerged: when fracture aperture exceeded 10 mm (Fig. 5), MRT increased substantially, ranging from 84 to 303 d (mean 172 d). This extended retention is primarily driven by soil infill within these larger fractures (porosity 0.41–0.74), which fundamentally transforms fractures from rapid downward conduits into highly retentive subsurface reservoirs. This structure-dependent storage mechanism provides a critical ecohydrological buffer, enabling infilled fractures to maintain a stable water supply under drought conditions (Hasenmueller et al., 2017; Zhang et al., 2016; Carrière et al., 2019; Yan et al., 2023; Howarth and Bishop, 2023).
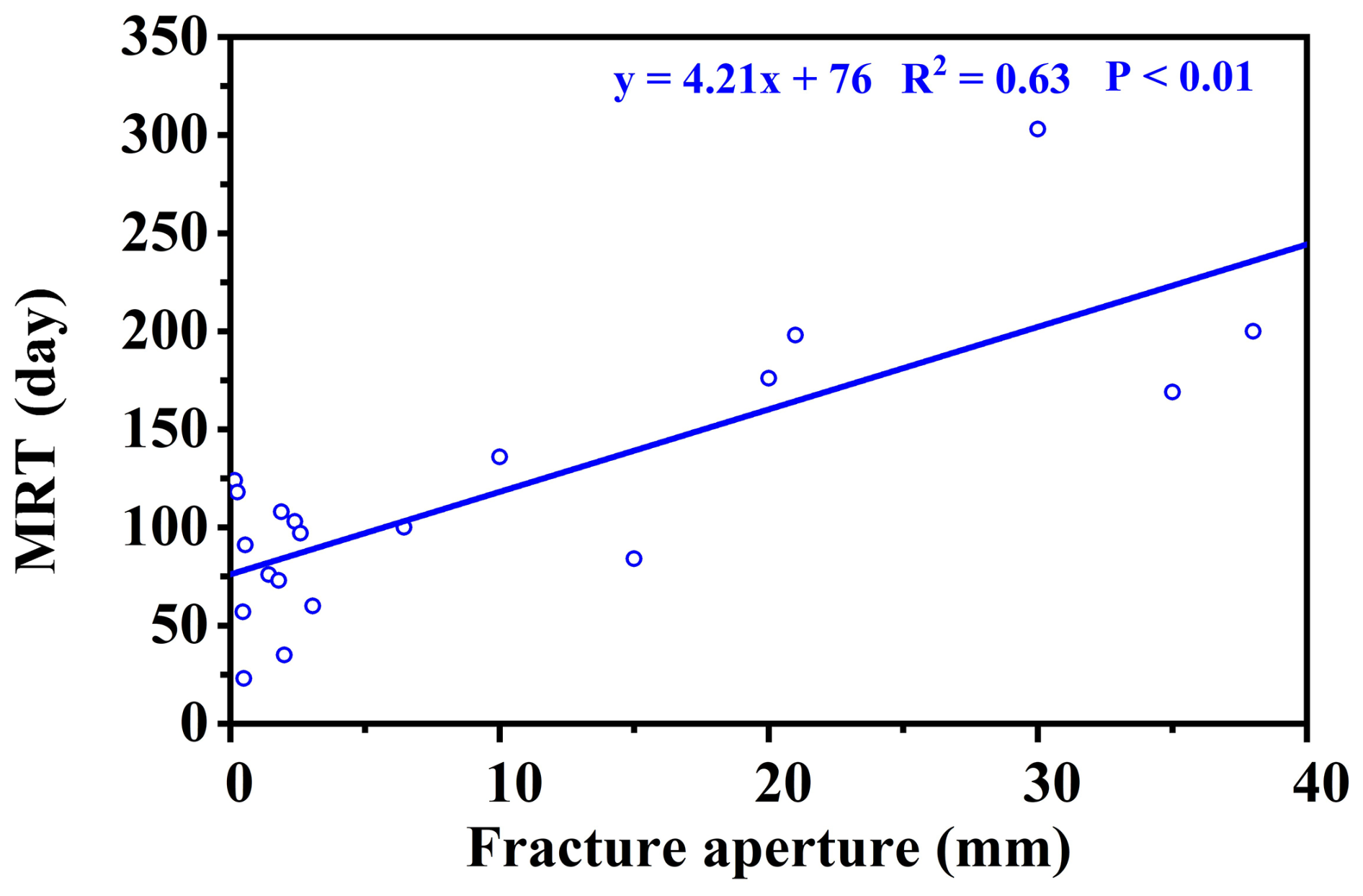
The distinct residence times of these environmental water pools directly shaped the water uptake strategies of different trees, as reflected in the MRT of their xylem water. (Note: xylem MRT characterizes the mixing volume and turnover time within the tree-source system, whereas MTT quantifies the transit lag time from source to xylem; Table C4). At site A, trees exhibited similar MRTs across all water sources (31–64 d), indicating a highly flexible, opportunistic strategy utilizing both transient mobile water and moderately retained rock water. In contrast, at sites with well-developed infilled fractures (Sites B and C and D) or thick retentive soils (Site E), trees demonstrated a targeted reliance on long-residence water to support sustained transpiration. For instance, the large tree ts6 at site C dynamically tapped into long-residence rock and bulk water (xylem MRT up to 124–128 d) to support its exceptionally high transpiration demand (average sap flow rate of 1.3 cm h−1). Similarly, evergreen trees (cc1–3) consistently utilized long-MRT bulk and rock water (73–76 d) to maintain year-round physiological activity compared to deciduous species. Furthermore, the interplay between soil retention and fracture traits dictates site-specific resilience. At site E, where rock fractures are poorly developed (apertures <2.6 mm) but soils possess high water-holding capacity (higher soil moisture content, Fig. B1c), the xylem MRT associated with bulk soil water (72–88 d) was notably longer than that of the limited rock water (51–61 d). This demonstrates that in the absence of large infilled fractures, the retentive soil matrix serves as the more stable and persistent moisture pool. Collectively, these xylem MRTs confirm that trees systematically align their root uptake with the specific geological compartments – either retentive soil matrix or infilled rock fractures – that offer the most stable, long-residence water supply.
3.3 Partitioning of plant water sources
Derived from the MixSIAR model outputs, Figs. 6 and D1 illustrate the proportional use of four water sources (bulk soil water, mobile soil water, rock fracture water, and infilled rock fracture water) across six phenological and hydrological periods (from senescence in October–November to late growing season in August–September). Our results reveal a clear structure-dependent water uptake strategy, where the importance of rock water is strongly governed by fracture traits and seasonal water availability, rather than uniform temporal shifts.
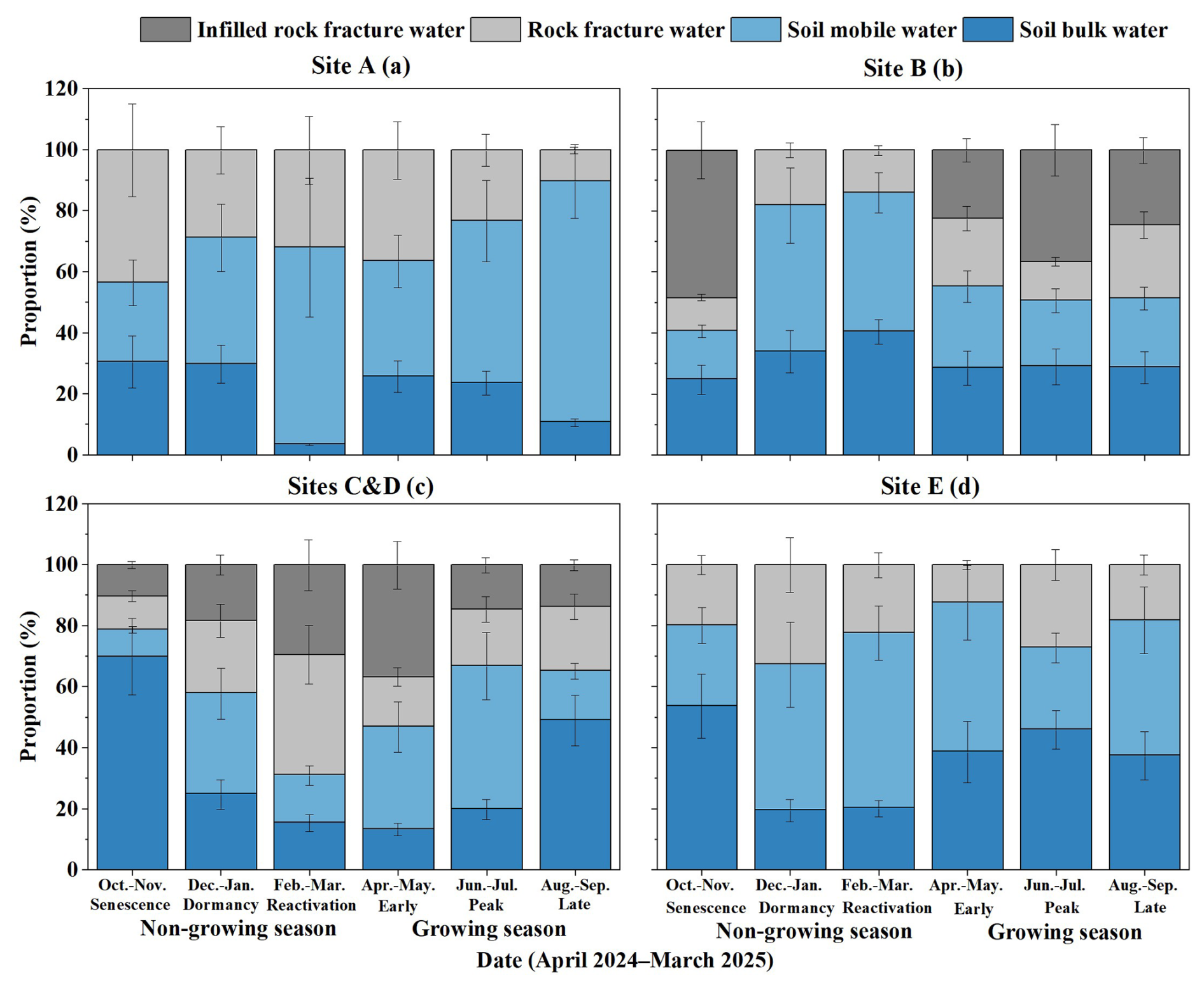
Figure 6Seasonal variations in the mean proportional contribution of water sources to tree xylem water, grouped by fracture aperture and infill characteristics. Data were collected from April 2024 to March 2025. The panels represent the representative fracture type defining each micro-habitat (noting that while minor fractures are ubiquitous, soil-filled fractures dominate storage at Sites B–D): (a) Site A: medium open fractures (aperture 3–10 mm, no infill); (b) Site B: fully infilled fractures (aperture 10–20 mm, fully infilled); (c) Sites C and D: wide partially infilled fractures (aperture >20 mm, partially infilled); (d) Site E: narrow open fractures (aperture <3 mm, no infill).The X axis displays data in 2 month intervals corresponding to distinct plant phenological stages (e.g., growing season (early growing, peak growing, late growing), senescence, reactivation, dormancy). Data represent the mean values of all sampled trees within each site group. Error bars indicate ±1 standard deviation (SD) derived from the MixSIAR model outputs.
During periods of abundant water supply, the influence of fracture properties on water sourcing was minimal. In the peak growing season (June–July; Figs. 6 and D1e), the contribution of mobile soil water increased markedly across all sites (mean 41 %), especially at sites A (mean 53 %) and C and D (mean 47 %). At sites C and D, the average contribution of infilled rock fracture water declined from 37 % (April–May) to 15 % (Fig. 6c), indicating that large fractures serve a regulatory storage function, storing excess water for later use, rather than acting as a dominant source during the rainy season.
Conversely, during dry and transitional periods (senescence, dormancy, reactivation, late and early growing season), tree water sourcing diverged significantly depending on local fracture aperture and infill conditions. At sites with highly developed and soil-filled fractures, trees demonstrated a strong capacity to exploit deep rock water to buffer drought. For instance, Table C3 shows that soil-filled fractures (>20 mm) at C and D have MRTs as long as 169–303 d (mean 212 d). These infilled structures act as stable, long-residence water pools that persist across seasons. As temperatures rose during the reactivation period (February–March), trees at sites C and D expanded their reliance on rock water to a mean of 69 % (Fig. 6c), with individual contributions ranging from 47 % to 88 % (Fig. D1c), supporting budburst before substantial rainfall resumed. This structural dependence persisted into the early growing season (April–May) and scaled with tree root-zone fracture volume. The largest tree (ts6, DBH 36 cm; Table A3), whose root zone contained the highest fracture volume (2.19m3, of which 87 %–96 % was soil-filled), derived up to 85 % of its water from rock sources, including 74 % from infilled rock fracture water (Fig. D1d). Additionally, Table C4 indicates that ts6 uses rock water with a residence time of up to 128 d, demonstrating its capacity to access stable, long-residence water for transpiration. Collectively, these findings confirm that large, infilled rock fractures function as critical “transitional reservoirs”, alleviating early-season soil water deficits.
This fracture-dependent buffering effect was also evident during the late-season dry-down. As rainfall and soil moisture declined in the late growing season and senescence periods, the availability of recent precipitation diminished. Trees at site B continued to rely heavily on infilled rock fracture water in senescence period (mean 49 % in Fig. 6b). Similarly, trees at site A (bp1–3) showed a substantial increase in rock fracture water use, jumping from mean 10 % in late growing season to mean 44 % in senescence period (Fig. 6a). Because large fractures maintain higher moisture content than rapidly depleting shallow soils (Fig. B2c vs. Fig. B1c), rock water provides an essential supplementary source preventing severe water stress during the wet-to-dry transition.
In stark contrast, trees at sites lacking large, infilled fractures exhibited entirely different survival strategies. Site E exhibited the least developed rock fractures (aperture Table C3). Throughout most of the year, including critical dry periods, these trees' reliance on rock water remained below 30 %. Without the deep buffering capacity of infilled fractures, these trees were forced to depend heavily on soil water. During the early growing season, individuals at site E relied on soil water for 82 %–91 % in Fig. D1d (mean 88 % in Fig. 6d) of their uptake. Even during the dormancy period (December–January), these rock-water-limited environments relied on minor rainfall pulses to recharge near-surface soil water (mean 67 % in Fig. 6d). However, due to the low rock permeability at this site (Kh<47 cm h−1; Table C3), water loss through deep percolation was limited. As a result, the average soil moisture content was relatively stable (Fig. B1c), enabling this soil-dependent strategy despite the absence of usable rock water.
4.1 Rock fracture properties drive isotopic variability and ecohydrological responses of vegetation
Our findings provide a critical karst-specific extension to the classic “ecohydrological separation” hypothesis. Traditionally, this concept posits that trees primarily utilize tightly bound soil water, while mobile water bypasses the root zone (McDonnell, 2014; Evaristo et al., 2015; Brooks et al., 2010). Conventional studies attribute this separation primarily to soil-specific factors, such as low soil water content, preferential flow paths, and coarse-textured, low-saturation soils with poor hydraulic conductivity (Finkenbiner et al., 2022, 2021; Liu et al., 2020; Sprenger et al., 2016). However, in karst landscapes characterized by extremely shallow soils, the soil matrix alone is insufficient to sustain year-round transpiration (Liu et al., 2025). Our results demonstrate that this functional separation is not confined to the soil but extends deep into the epikarst architecture. Crucially, the structural complexity of the karst subsurface – specifically variations in fracture aperture and the degree of soil infill – creates a massive, compartmentalized reservoir of “bound” water, limiting mixing among water pools and reinforcing their functional separation.
Our study reveals significant isotopic variability in fracture water across different depths and fracture apertures (Fig. B4a and b). In the 0–200 cm depth range, the Lc-excess values of fracture water are close to 0 ‰, with many values even falling below 0 ‰, reflecting the influence of recent rainfall inputs coupled with kinetic fractionation due to near-surface evaporation.. Conversely, deep fracture water at depths greater than 200 cm typically has Lc-excess values above 0 ‰. These signatures suggest that deeper fracture water is largely decoupled from surface evaporative processes and is likely derived from infiltration events that preserve the original isotopic composition of precipitation. More importantly, our data reveal a clear structure-dependent threshold behavior governing water availability. Fracture aperture further influences the isotopic characteristics of stored water. Fracture water within narrow apertures (0.5–2 mm) displays a wide range of δD and δ18O values that exceed the isotopic variability observed in plant xylem water. This variability indicates that water stored in small fractures is more susceptible to mixing processes and variable evaporative fractionation. In contrast, fracture water within larger apertures (10–38 mm) falls largely within the isotopic range of xylem water and shows Lc-excess values closer to those of plant water. These patterns indicate that larger fractures, particularly those containing soil infill, can reduce evaporative enrichment and slow water exchange with the surrounding matrix. Previous work has shown that soil infill can substantially modify fracture hydraulic properties(Liu et al., 2024c), increasing water retention and creating relatively stable subsurface water stores that may support plant water uptake.
Fracture water at 600 cm depth shows more concentrated δD and δ18O values, with a much narrower isotopic range than xylem water. Therefore, if this deep fracture water is used to represent root zone fracture water for vegetation water source partitioning, it may fail to explain the entire water utilization strategy of vegetation and lead to errors. Many studies that use deep spring water (Ding et al., 2021; Rong et al., 2011; Fan et al., 2023; Deng et al., 2015; Nie et al., 2012; Cai et al., 2023, 2025; Wu et al., 2021, 2024b; Carrière et al., 2019; Zeng et al., 2021), or mixed borehole water (Deng et al., 2020) as epikarst zone water may introduce uncertainty in identifying the actual water sources used by plants.
Our findings suggest that analyses of plant water use in karst systems should explicitly consider fracture structural properties (e.g., aperture, depth, and infill porosity) when defining rock water sources, in order to better resolve water partitioning within the coupled soil-plant-rock fracture continuum.
4.2 On the surprising similarity between mobile soil water and bulk soil water
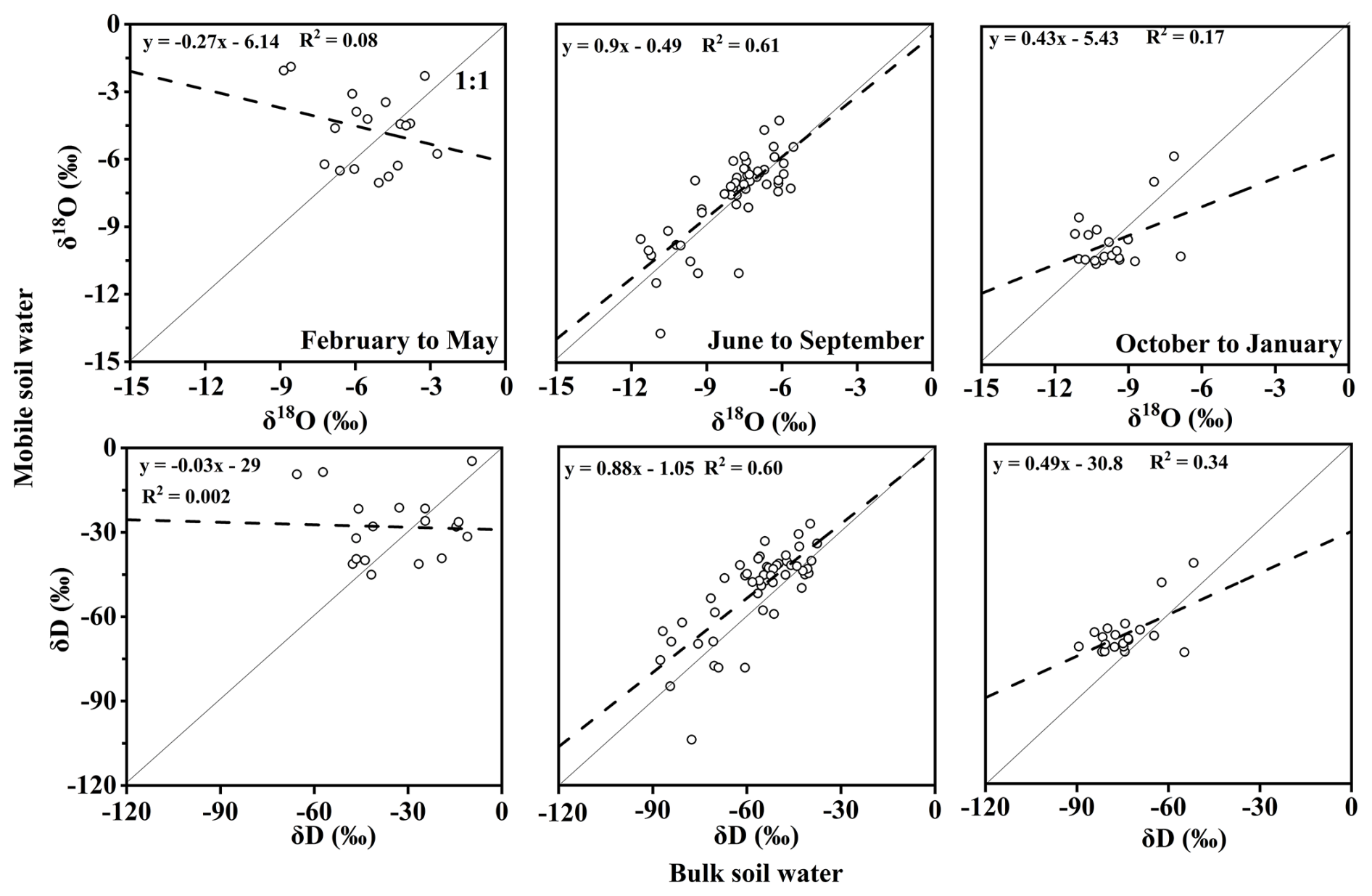
Our isotopic correlations between bulk soil water and mobile soil water (Fig. B5) show that during dry seasons (February–May and October–January), the correlation between the isotopic values of bulk soil water and mobile water is weak (R2=0.002–0.34). This indicates poor hydraulic coupling between the two components. Notably, the isotopic values of bulk water (obtained using the CVD method) are more depleted than those of mobile water (using suction lysimeters). This is consistent with the initial findings of Brooks et al. (2010) and follow-up work by Sprenger et al. (2019), Zhao and Wang (2021) and Xu et al. (2025). Brooks et al. (2010) conducted their study in a Mediterranean climate within the western Cascade Mountains of Oregon, USA. In the H. J. Andrews Experimental Forest, they collected samples during the dry season (June, August, September 2004–2005) and the wet-up period in autumn 2006 (October–December). Across these campaigns, Brooks et al. (2010) found that the weak isotopic correlation between bulk and mobile water suggests limited mixing, likely resulting from their residence in different pore domains. This implies that during these periods, bulk and mobile water are influenced by distinct recharge sources and subject to different degrees of evaporation-driven isotopic fractionation, thereby enhancing ecohydrological separation.
However, in our study, we observed that during the high-rainfall period in June–September, this separation effect becomes less pronounced. The correlation between bulk and mobile soil water isotopic values improves markedly (R2=0.60–0.61; Fig. B5), indicating that recent precipitation inputs increasingly dominate the mobile water pool and that some degree of hydraulic exchange occurs between the two compartments. This seasonal shift suggests a dynamic and partial coupling between soil water pools under high moisture conditions, which may temporarily weaken ecohydrological separation (Geris et al., 2015). Snelgrove et al. (2021) report that in a catchment in northern Britain, high soil water storage and limited precipitation inputs in humid settings can facilitate mixing between soil water and mobile water, reducing the likelihood of pronounced separation. Similarly, in ecosystems with high rainfall and elevated soil moisture, such as tropical rainforests, the isotopic differences between soil water and plant xylem water are often small or even negligible, indicating little to no separation (Liu et al., 2020). Ecohydrological separation is more likely during dry periods and tends to weaken or disappear during the wet season (Luo et al., 2019; Hervé-Fernández et al., 2016).
To further investigate the hydraulic linkage of different water components at varying depths, we analyze isotopic relationships stratified by depth. The results show that in the upper 0–50 cm layer, the correlation between soil bulk and mobile water is relatively strong (R2=0.68–0.78), whereas in the 50–200 cm layer, the correlation weakens significantly (R2=0.32–0.46; Fig. 7). Vargas et al. (2017) demonstrated that under wet conditions, 75 %–95 % of mobile water can exchange isotopically with surrounding bound water in the upper soil layers (<30 cm).
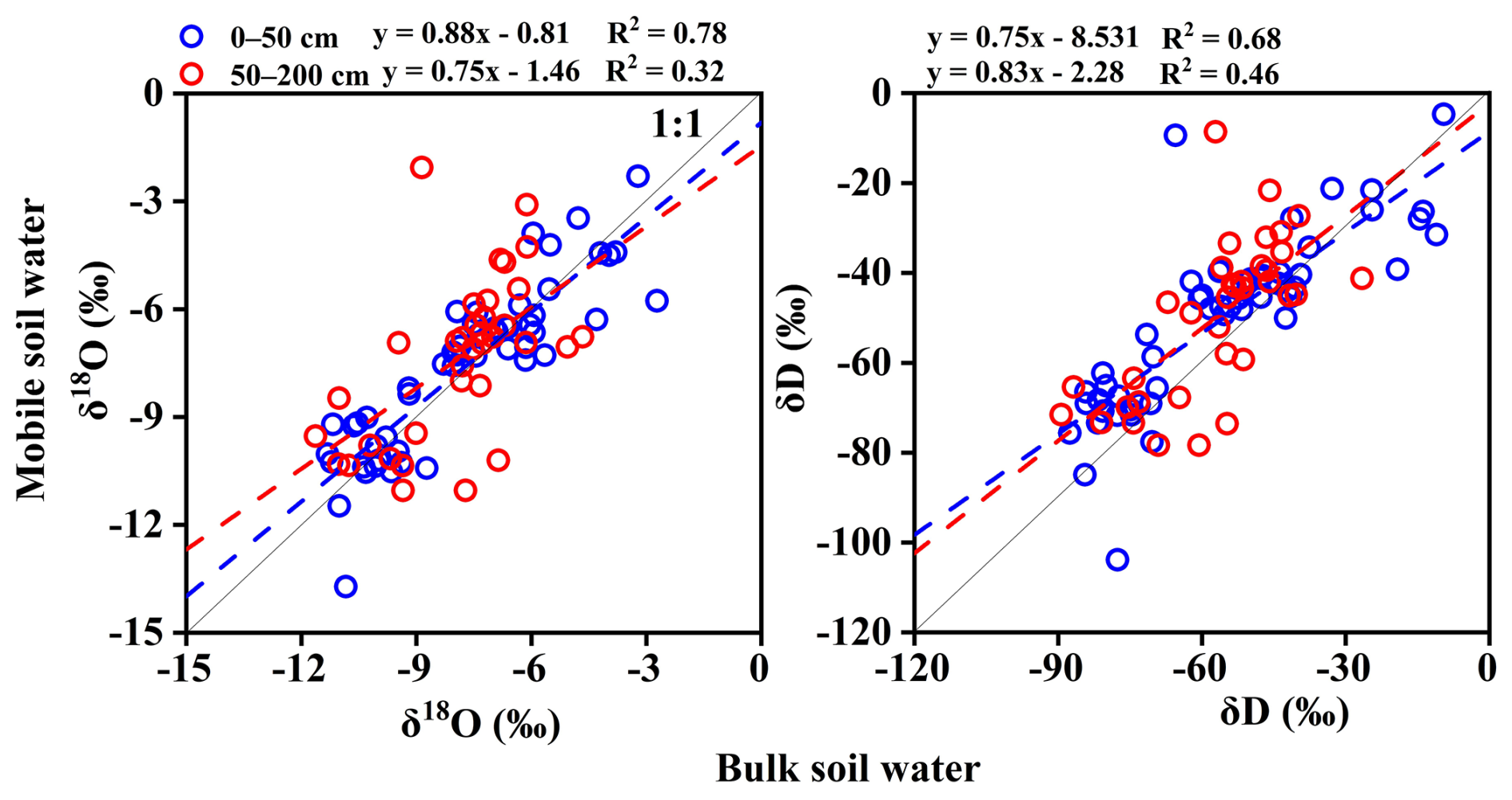
Figure 7Isotopic comparisons (δD and δ18O) between bulk soil water and mobile water across soil depths.
Figure B6 further reveals that with increasing depth, mobile soil water becomes increasingly enriched in δD and δ18O, while its isotopic range remains relatively stable, and Lc-excess values remain near 0 ‰. In contrast, bulk soil water shows a gradual depletion in δD and δ18O with depth, along with significantly lower Lc-excess values, aligning more closely with xylem water.
These findings suggest that in surface soils, precipitation infiltrates rapidly and alters the isotopic composition of both mobile and bulk soil water, while also enhancing hydraulic connectivity between different pore domains. This is particularly evident during periods of intense rainfall, when high soil moisture content reduces matric potential gradients and facilitates lateral and vertical water exchange across pore domains. Under such conditions, even tightly bound water may undergo partial displacement or mixing due to sustained infiltration and elevated hydraulic gradients.
With increasing depth, however, the hydraulic connection between bulk and mobile water progressively weakens. Deep bulk water increasingly behaves as a relatively isolated reservoir, with limited exchange, while mobile water remains hydrologically dynamic and responsive to recent precipitation inputs through preferential flow pathways. This vertical stratification in hydraulic behavior results in the clear manifestation of ecohydrological separation within the soil profile under most seasonal conditions.
Nevertheless, during the peak rainy season, the high volume and intensity of precipitation enhance water fluxes and recharge rates throughout the profile. This leads to greater pore connectivity and transient mixing between bulk and mobile water pools, thereby weakening the separation effect. Thus, our findings demonstrate that ecohydrological separation is not static but modulated by seasonal hydrological regimes, with stronger separation under drier conditions and partial coupling during wetter periods when hydrodynamic forces override structural constraints.
4.3 A perceptual model for the role of rock fractures in plant water uptake in karst
Our findings lead to a perceptual model that illustrates how plants dynamically access different water sources in karst ecosystems, shaped by the structural and hydrodynamic properties of the soil–rock continuum. The conceptual framework presented in Fig. 8 depicts a vertically stratified water source system composed of four primary compartments: mobile soil water, bulk soil water, rock fracture water, and infilled rock fracture water.
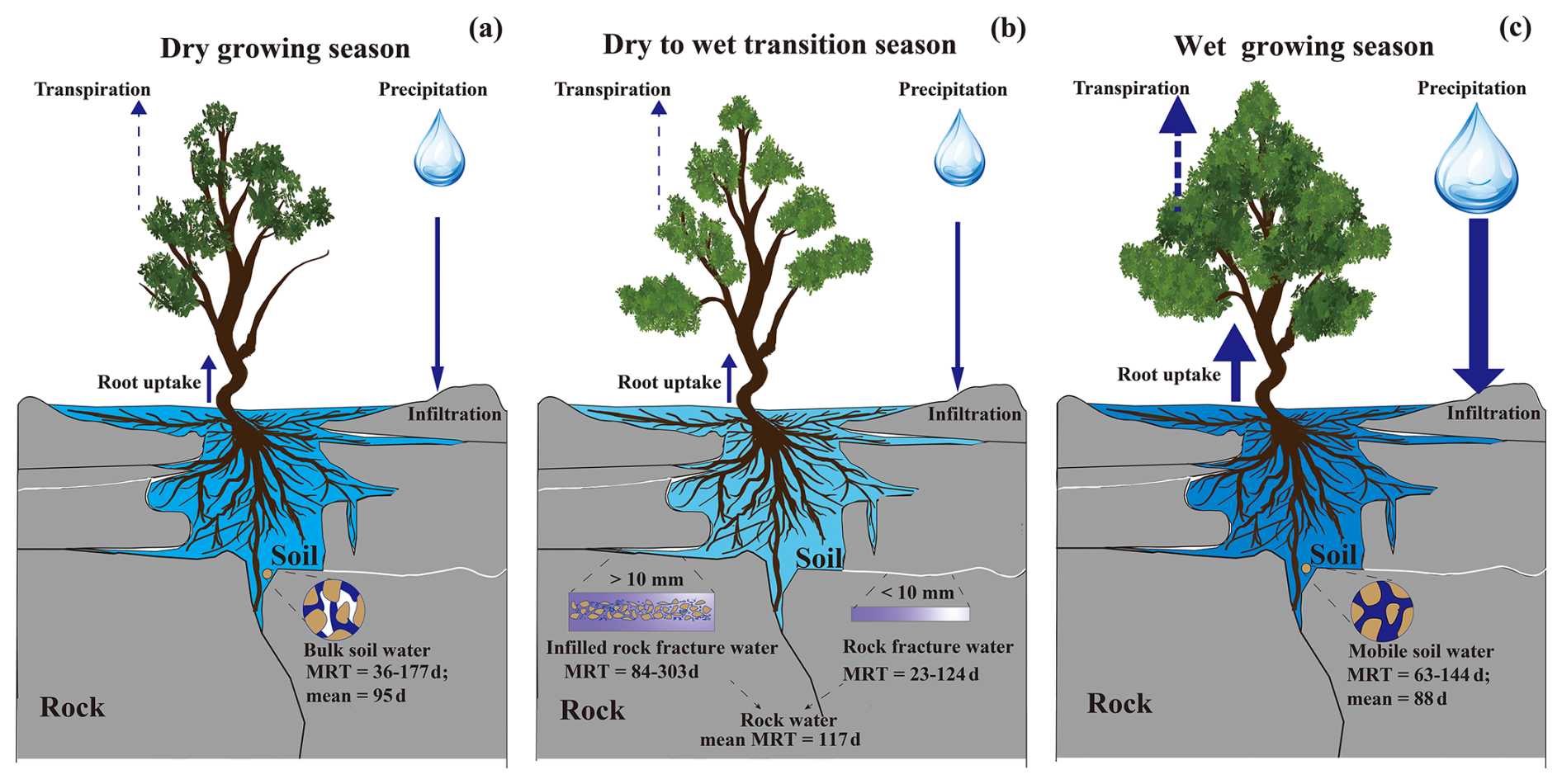
Overall, the preferential water source hierarchy in this karst system follows the order: mobile soil water > bulk soil water > rock water. In our study, mobile water is characterized by short residence times and high turnover. Mobile soil water is the primary support for transpiration during wet periods but is less accessible during dry spells. In contrast, bulk soil water, with its longer MRT, is mainly used by vegetation during the late growing season and senescence period. Rock water, especially that stored in soil-filled fractures, with its longer MRT and higher retention capacities, acts as a more stable source, sustaining plant function when soil water is depleted, thereby substantially enhancing tree survival and transpiration stability during drought periods and facilitating the onset of the growing season (Hahm et al., 2022; Leite et al., 2025; Schwinning, 2020).
Specifically, precipitation infiltrates the thin soil layer and preferentially recharges mobile soil water stored in connected pores. This water is readily available for root uptake but has a short residence time and low retention capacity, particularly during the wet season (peak growing season). As water percolates downward, part of it is retained in micropores as bulk soil water, which is replenished more slowly and supports vegetation during late growing season and senescence period. At greater depths, plant roots interact with the weathered bedrock. Rock fractures with smaller apertures and minimal soil infill function as transient water storage compartments with intermediate MRT, whereas larger fractures that are substantially filled with soil serve as long-term reservoirs, characterized by significantly prolonged water retention (MRT up to 303 d in our study). These fractures are crucial for maintaining transpiration during dry or transitional periods (e.g., reactivation period and early growing season), especially for large, deep-rooted trees. Our isotope and MRT analyses show that vegetation flexibly shifts water use based on seasonal availability, progressively relying on infilled rock fracture water and bulk soil water as mobile soil water becomes scarce.
In Southwest China, long-term anthropogenic disturbances and natural processes have driven extensive karst desertification, resulting in severe soil degradation, vegetation loss, and ecosystem instability (Wang et al., 2019b). Despite a general “greening” trend since 2000 (Tong et al., 2018), projections indicate that drought frequency in Southwest China is likely to increase further between 2021 and 2035 (Compilation Committee of the Assessment Report on Climate Change in Southwest China, 2021). A combination of reduced precipitation, rising temperatures, more frequent and intense droughts, soil degradation, and insufficient water resource management may amplify the vulnerability of current forest restoration efforts (Peng et al., 2020; Zhou et al., 2020; Lin et al., 2015; Zhao et al., 2024), affecting plant water uptake and long-term ecosystem stability. With global warming intensifying the hydrological cycle, shifts between wet and dry periods are becoming more pronounced (He et al., 2024; Chen et al., 2025). It is projected that by 2040–2100, nearly 60 % of the global land surface will experience accelerated dry-wet transitions (Chen and Wang, 2022). The frequency of both atmospheric drought and surface soil drought is rising (Xu et al., 2024; Fabiani et al., 2024), which may lead to fundamental changes in plant water-use strategies (Wei et al., 2023; Salomón et al., 2022). Under such conditions, rock fracture water may serve as a crucial buffer, sustaining vegetation during drought events (Schwinning, 2020; Korboulewsky et al., 2020; Luo et al., 2024b; Wu et al., 2024a). However, current research has not adequately elucidated the regulatory mechanisms by which soil–rock structures control water transport and retention times, nor their influence on vegetation water uptake. In particular, there is a lack of quantitative, mechanistic studies focusing on factors such as fracture aperture, soil-filled fractures, soil physical properties, and the interactions between vegetation and the soil–rock structure.
Overall, our framework underscores the critical, yet often underappreciated, role of rock fracture water as a hydro-buffer that sustains vegetation function amidst increasing climate variability in karst regions.
4.4 Study limitations and future research directions
The study design incorporates comparisons of identical tree species across sites and different species within similar structural settings, allowing partial separation of structural and biological controls. Representative fracture types were also sampled repeatedly within the study area. However, the results are derived from a single lithological setting (limestone) in a subtropical karst region and should therefore be interpreted as a process-based case study. Differences in plant water-use strategies between limestone and dolomite habitats have been reported (Liu et al., 2026; Ding et al., 2025), suggesting that lithological controls may influence the generality of the observed patterns. The extent to which these findings are applicable across different rocky habitats, lithologies, and climatic regimes remains to be evaluated.
Uncertainty also arises from the estimation of MRT, which is based on fitting seasonal isotope signals and assumes periodic input variability. In heterogeneous subsurface systems, deviations from these assumptions can affect residence time estimates, and such metrics are therefore more appropriately interpreted as effective measures of storage (Kirchner, 2016; Benettin et al., 2017).
Although uncertainty in isotope-based source partitioning has been reduced by directly sampling individual fractures, some uncertainty remains due to partial overlap among endmembers and the structural heterogeneity of subsurface water pools. This limitation has been widely discussed in isotope-based mixing approaches and ecohydrological separation studies (Evaristo et al., 2015; Sprenger et al., 2019). In addition, representing fracture water as discrete categories simplifies the continuous variability of fracture networks in terms of aperture, connectivity, and storage properties (Hartmann et al., 2014).
Future research should extend this approach across a broader range of lithologies and climatic settings to test the generality of fracture-controlled water uptake. Fracture-scale sampling in diverse rock environments will help clarify how structural properties regulate plant water use. Integrating isotope-based methods with direct observations of root distribution within fractures, and explicitly incorporating fracture structural parameters into soil–rock continuum ecohydrological models, will improve the representation of water storage and uptake in bedrock systems and support more effective ecosystem resilience and karst forest restoration strategies.
Our study represents one of the first mechanistic investigations into the role of rock fractures in facilitating rock moisture uptake by plants in karst environments. By integrating stable isotope analysis with MRT modeling, we delineated four functionally distinct water compartments within a structurally heterogeneous soil–rock continuum: mobile soil water, bulk soil water, rock fracture water, and infilled rock fracture water. Our results demonstrate that plant water uptake strategies are seasonally modulated by the availability and hydrodynamic behavior of these compartments. Notably, vegetation exhibits a clear preference for water sources with shorter MRTs, especially under wetter conditions. During the peak growing season, rainfall-driven mobile soil water predominates in supporting plant transpiration. In contrast, during no growing or transitional seasons, trees increasingly rely on bulk soil water and rock fracture water, particularly from infilled fractures characterized by longer MRTs (84–303 d). The limited mixing and distinct MRTs observed among these water pools are largely consistent with the premise of the ecohydrological separation hypothesis. However, under high moisture conditions, increased hydraulic connectivity appears to reduce this separation. The perceptual model developed from this work highlights the critical buffering role of rock fracture water in sustaining vegetation under variable hydroclimatic regimes. These findings underscore the importance of incorporating bedrock heterogeneity, water source dynamics, and seasonal shifts in plant water use into ecohydrological models of karst systems. Given the anticipated increases in drought frequency and climatic variability, future forest management and restoration strategies in karst landscapes should explicitly consider the integrated behavior of the soil–rock-plant continuum to enhance ecosystem resilience.
Table A1Summary of soil moisture monitoring and soil water sampling design across study sites.
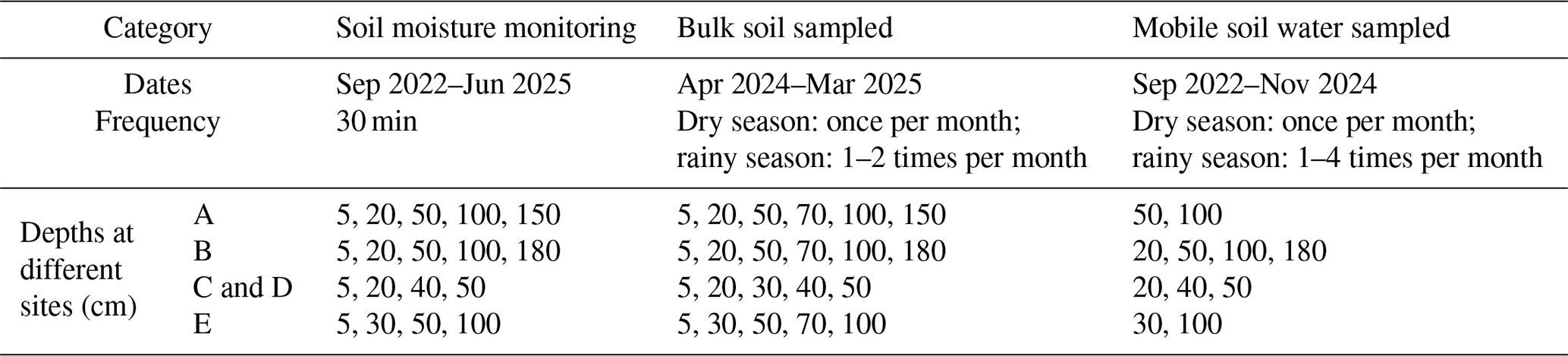
Table A2Summary of rock moisture monitoring and rock water sampling design across study sites.
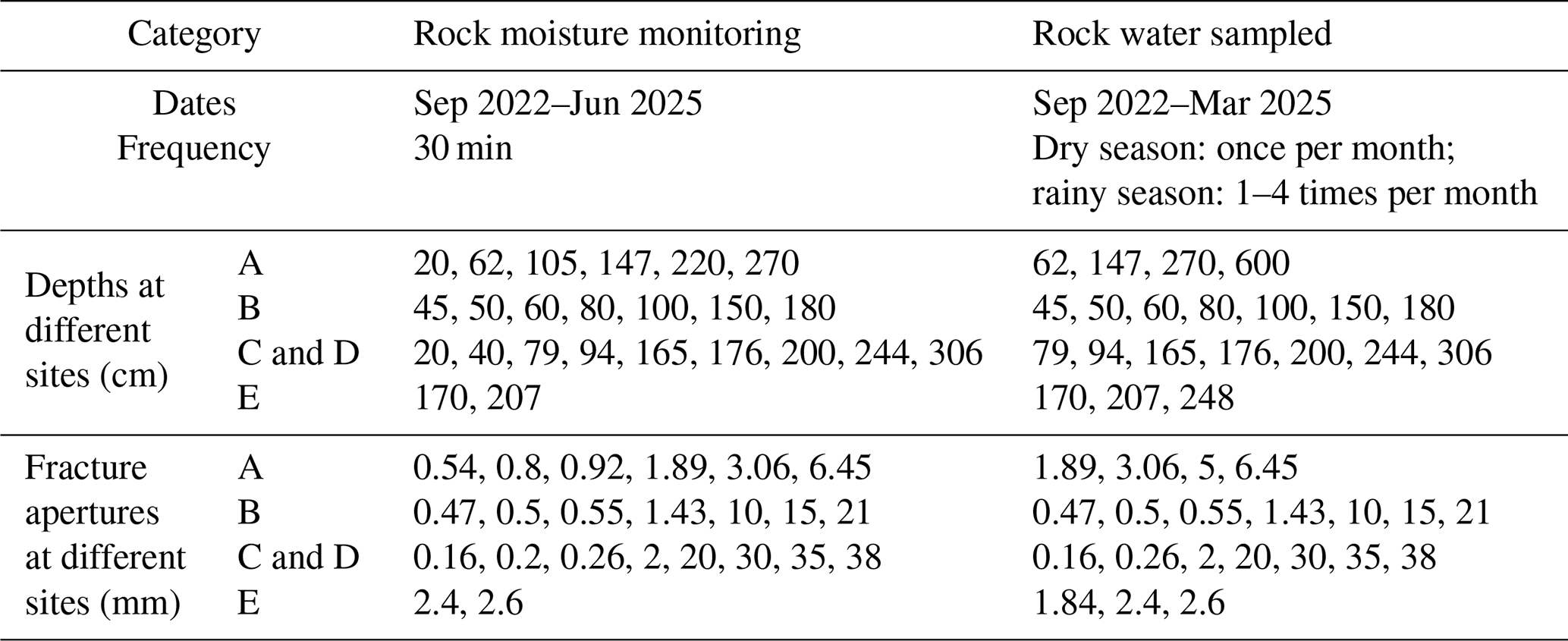
Table A3Summary of tree-based sampling design and root-zone characteristics across study sites.
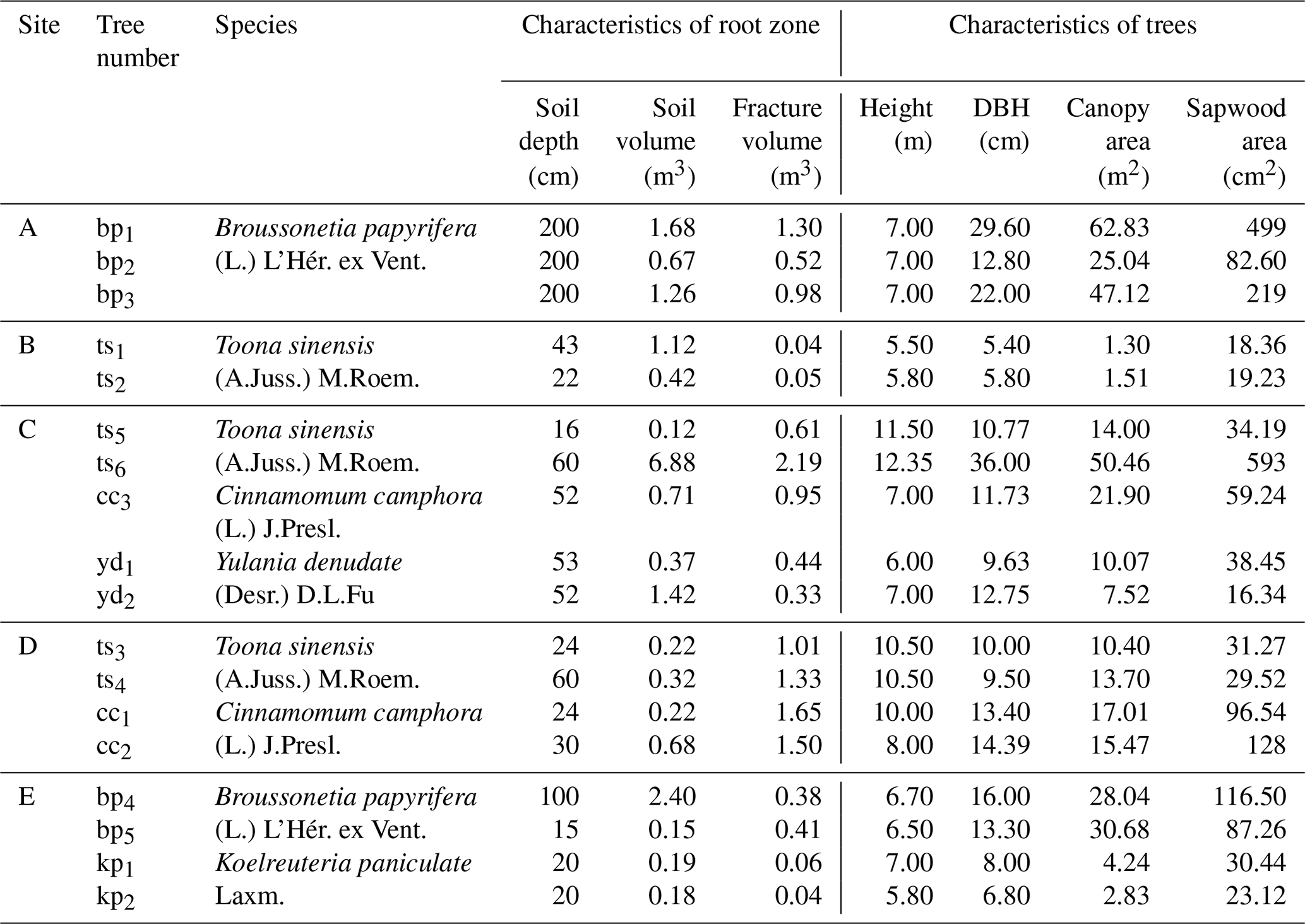

Figure A1Field photographs illustrating soil and rock water sampling procedures. (a) Vertical boreholes drilled at sites C, D, and E with water sampling devices installed at predefined depths. (b) Horizontal boreholes drilled into exposed bedrock outcrops at sites A and B with sampling devices installed within the soil–rock profile. (c) Collection of soil samples for laboratory extraction of bulk soil water. (d) Collection of mobile soil water using suction lysimeters installed in soil layers. (e) Collection of rock fracture water from narrow fractures (aperture approximately 0.1–10 mm) with little or no soil infill. (f) Collection of infilled rock fracture water from wider fractures (>10 mm aperture) largely filled with soil or fine sediments.
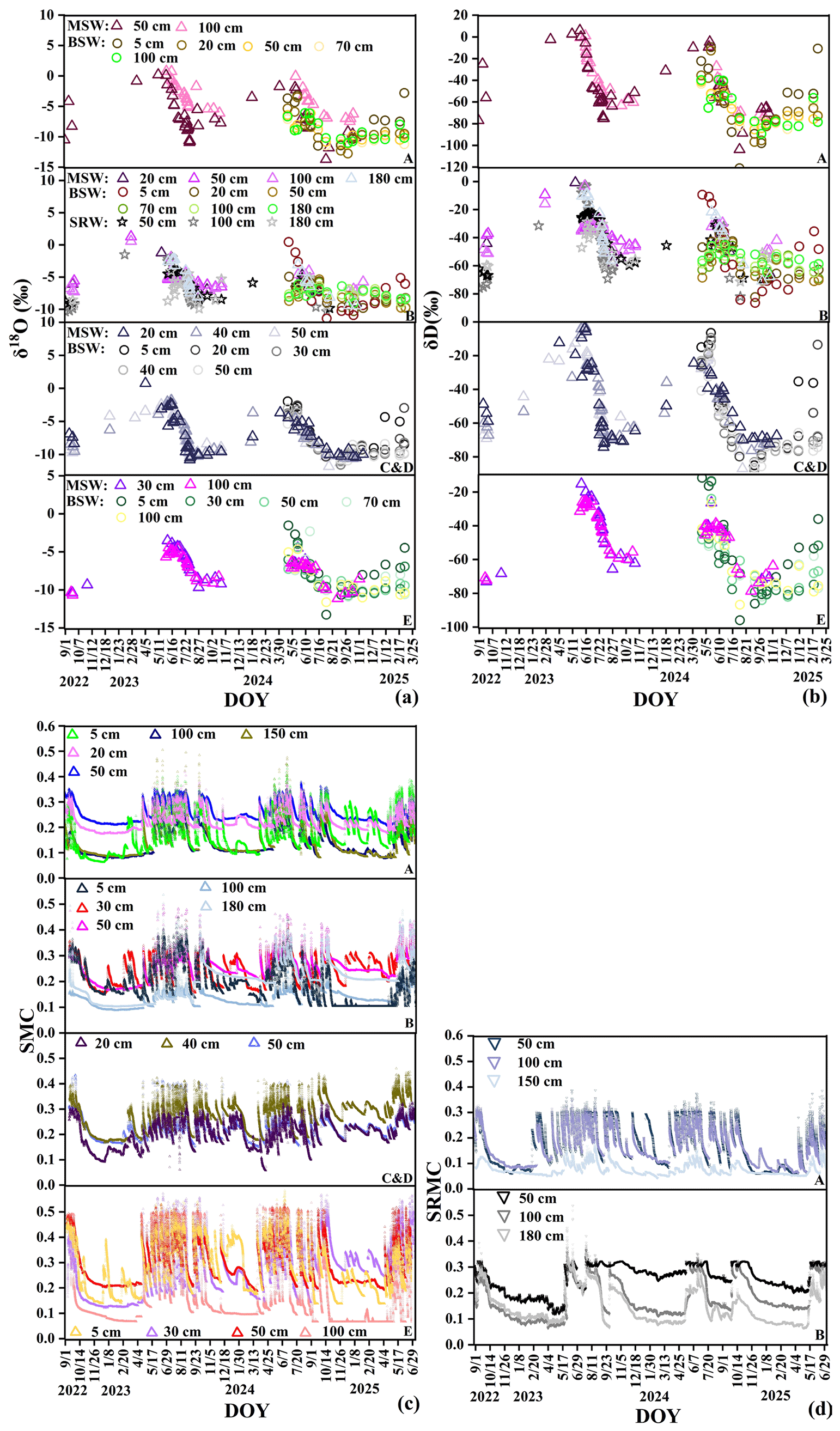
Figure B1Dynamic variations in stable isotopes of soil water at different depths at sites (A–E): (a) δ18O and (b) δD, and moisture content: (c) and (d). Mobile soil water (MSW), bulk soil water (BSW), soil–rock interface water (SRW), soil moisture content (SMC), soil–rock interface moisture content (SRMC).
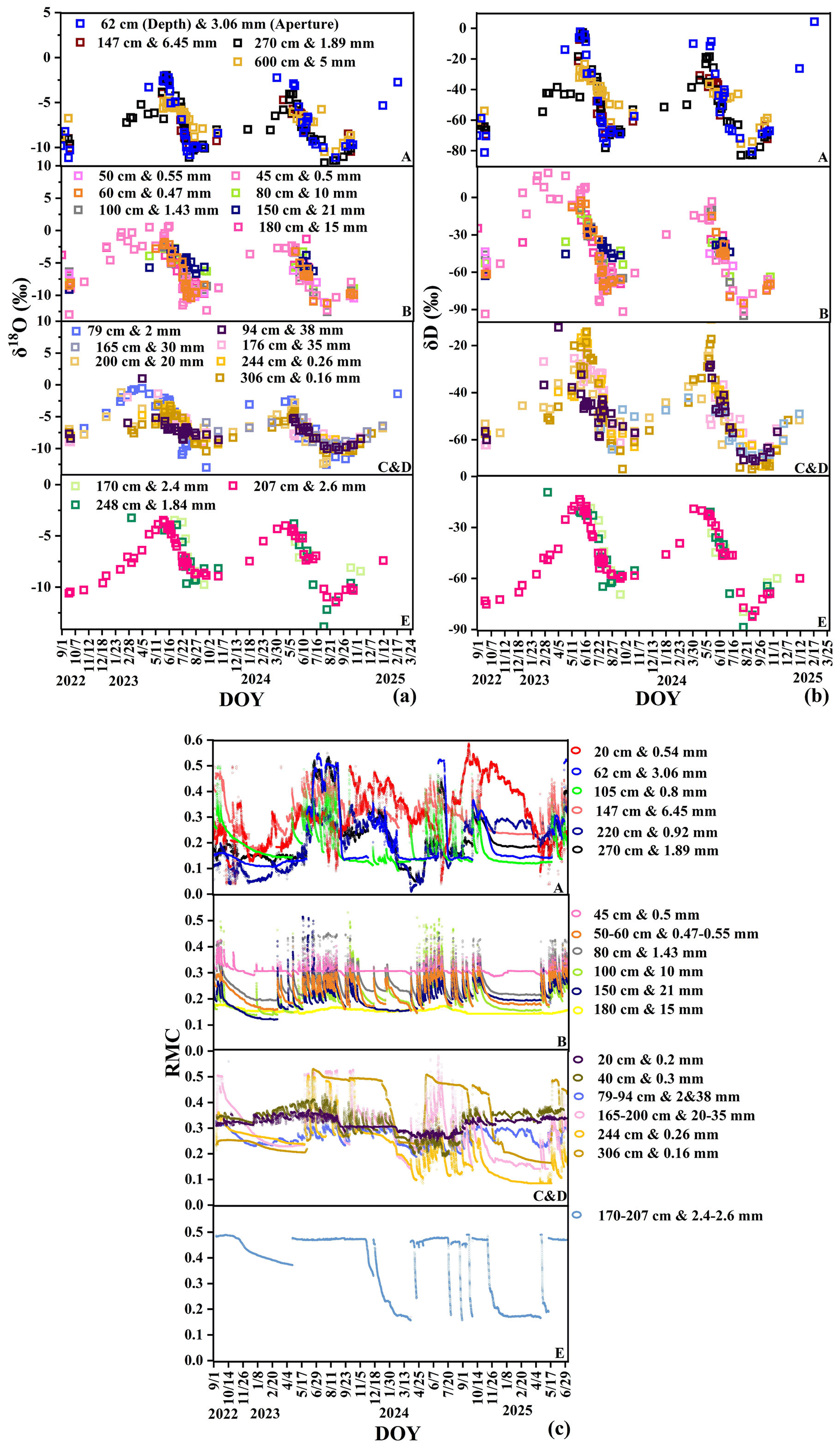
Figure B2Dynamic variations in stable isotopes of rock fracture water at different depths/apertures at sites (A–E): (a) δ18O and (b) δD, and moisture content: (c).
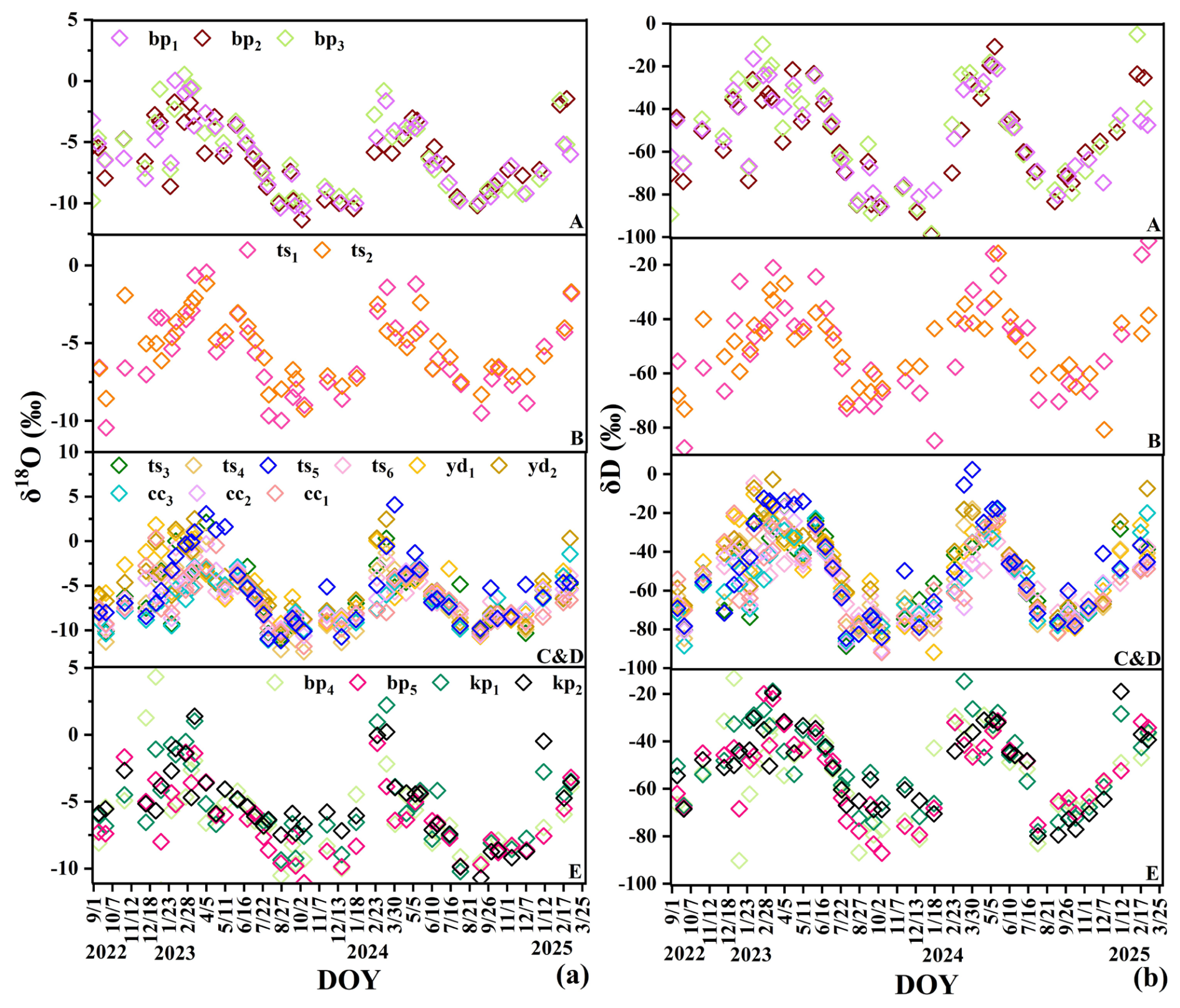
Figure B3Dynamic variations in stable isotopes of xylem water (δD and δ18O) in different trees at sites (A–E).
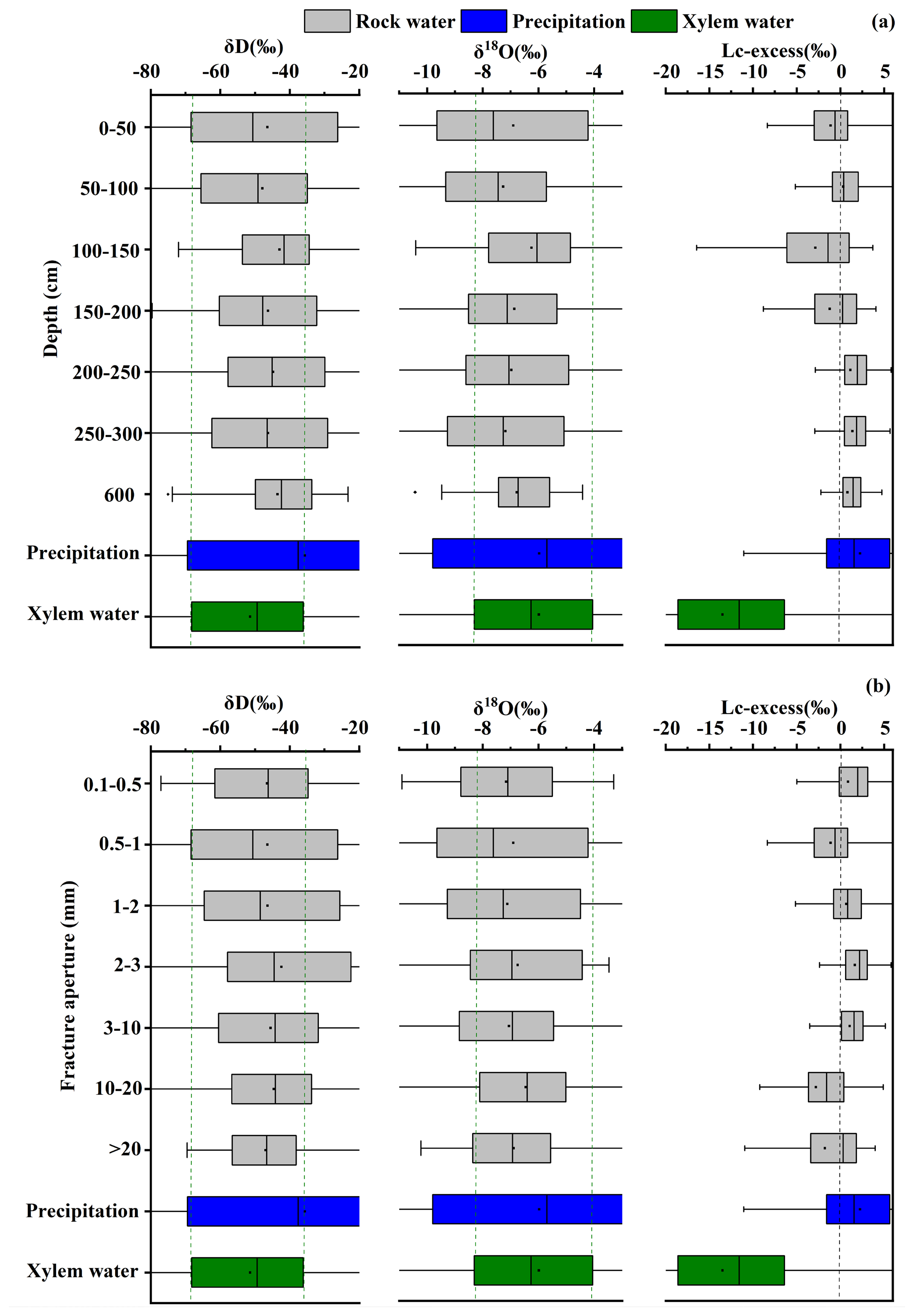
Figure B4Average values and ranges of δD, δ18O and Lc-excess in rock waters with different apertures.
To distinguish it from mean residence time (MRT), we applied the mean transit time (MTT) phase shift method from Allen et al. (2018) to quantify the time delay in water transport between compartments. The phase shift primarily captures the time lag between input signals (e.g., precipitation isotopic composition) and output signals (e.g., plant or groundwater isotopic composition). MTT represents the time lag or transmission time between different compartments in the hydrological cycle (e.g., precipitation to soil, soil to rock, rock to plant).
The MTT (in day) calculation was made by:
Where is the angular frequency constant, which represents the yearly cycle (assuming a one-year period for seasonal variations); ϕp is the phases of precipitation sine curves, and ϕw is water sine curve (mobile soil water, bulk soil water, rock fracture water, infilled rock fracture water, soil–rock interface water and xylem water).
Table C1 shows MTT was positively correlated with MRT (MTT , R2=0.77), with a range of 6–82 d and an average of 28 d. Site B also exhibited significantly higher MTT values (10–82 d, mean 39 d) compared to other sites (638 d, mean 21 d). Data in Tables C1–C3 consistently show that MRT exceeds MTT. This disparity reflects that the two metrics describe different aspects of water age dynamics within the same compartment. MTT is derived from the phase shift of the seasonal isotope signal and therefore represents the timing of signal propagation between connected compartments. In contrast, MRT is derived from amplitude damping and reflects the persistence of water within a reservoir as controlled by storage volume and internal mixing. Because subsurface storage and mixing can substantially attenuate isotope amplitudes without proportionally delaying signal transmission (due to preferential flow pathways), MRT is typically longer than MTT. Despite this difference in magnitude, the positive relationship between MRT and MTT indicates that subsurface structural properties coherently influence both transport delay and storage persistence.
Table C4 details both the MRT and MTT for different water sources utilized by trees. Here, MRT (derived from amplitude damping) characterizes the mixing volume and turnover time of water within the tree-source system (storage), while MTT (derived from phase shift) quantifies the time lag for water to be transported from the soil/rock source to the tree xylem (uptake delay). Negative MTT values occur when the isotope signal in plant xylem precedes that of water sources. This does not indicate negative physical transport time, but rather reflects the dominance of older or deeper water pools in plant uptake, where isotope signals are seasonally advanced due to prior recharge or long-term storage (Liu et al., 2024d).
Table C1Statistics of MRT and MTT (day) for mobile and bulk water in different soil layers.
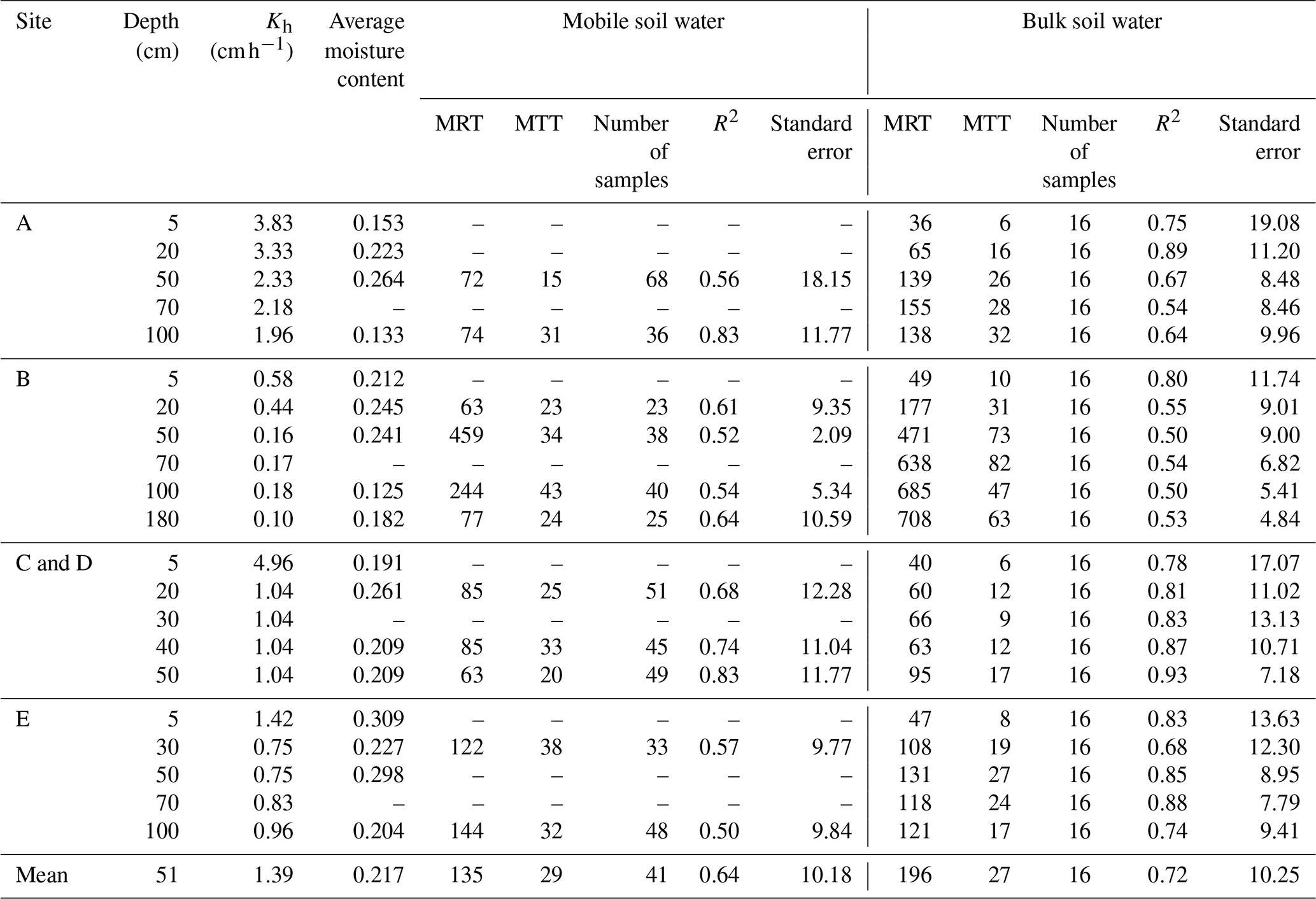
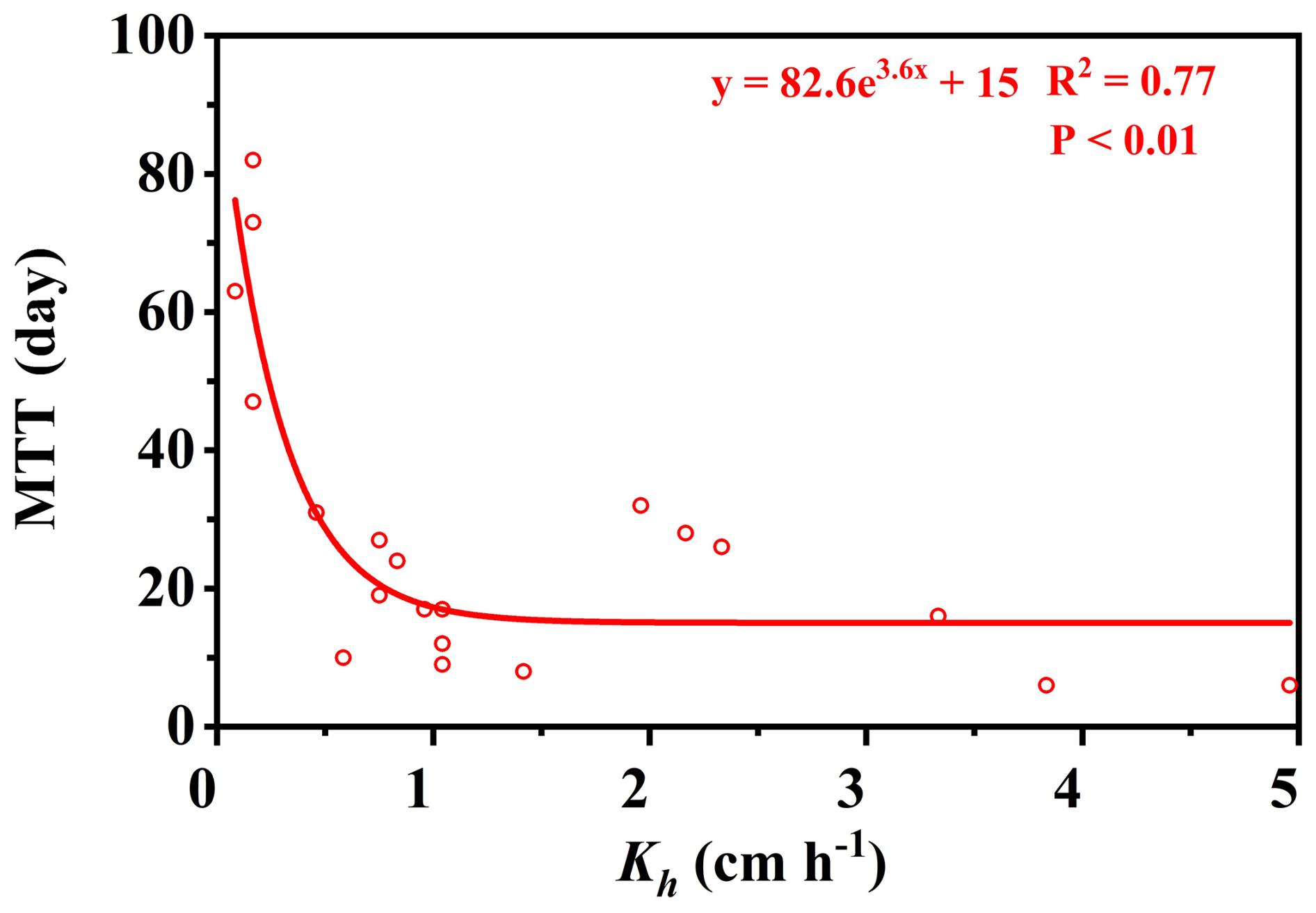
Table C2Statistics of MRT and MTT (day) for soil–rock interface water in different soil layers.
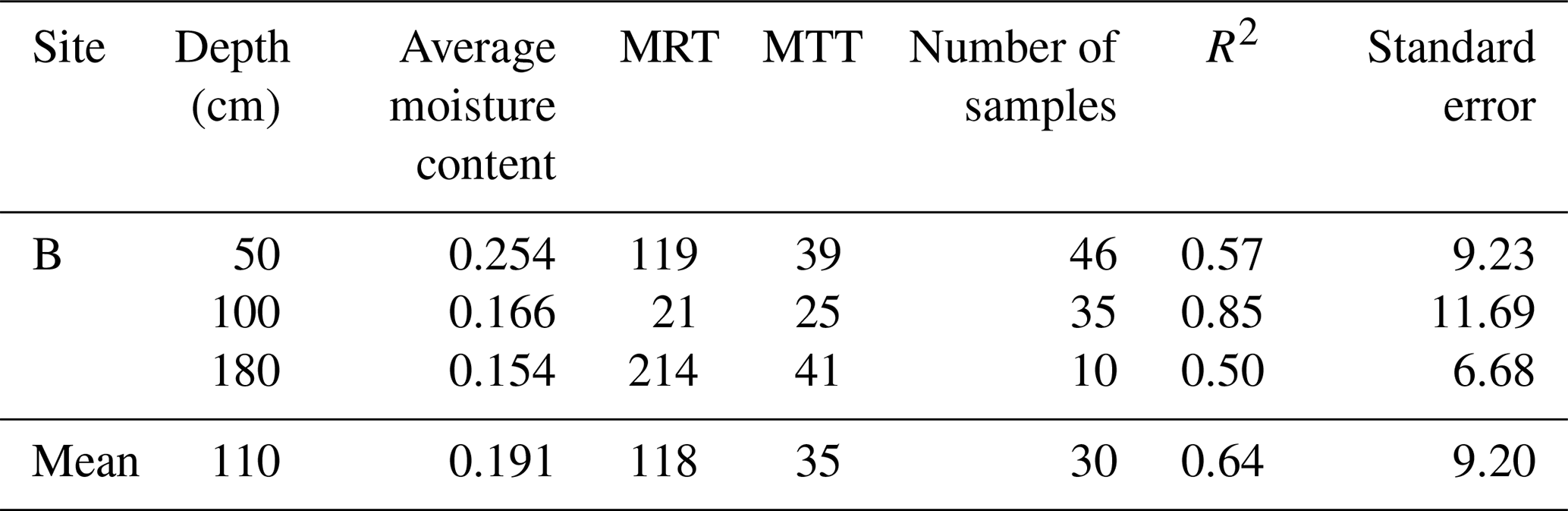
Table C3Statistics of MRT and MTT (day) for fractures of different apertures and depths.
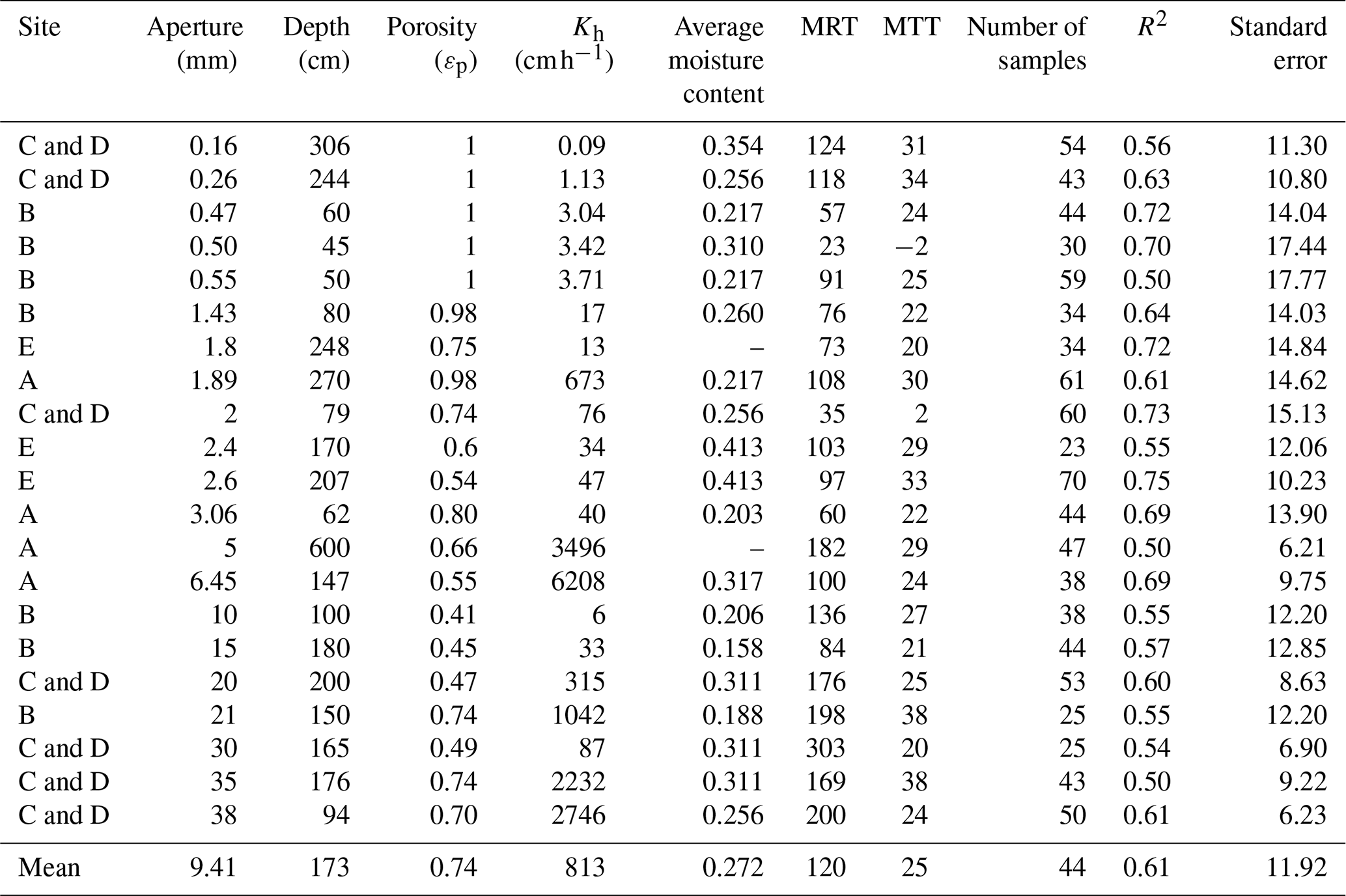
Table C4Statistics of MRT and MTT (day) for different trees at typical sites in the study area.
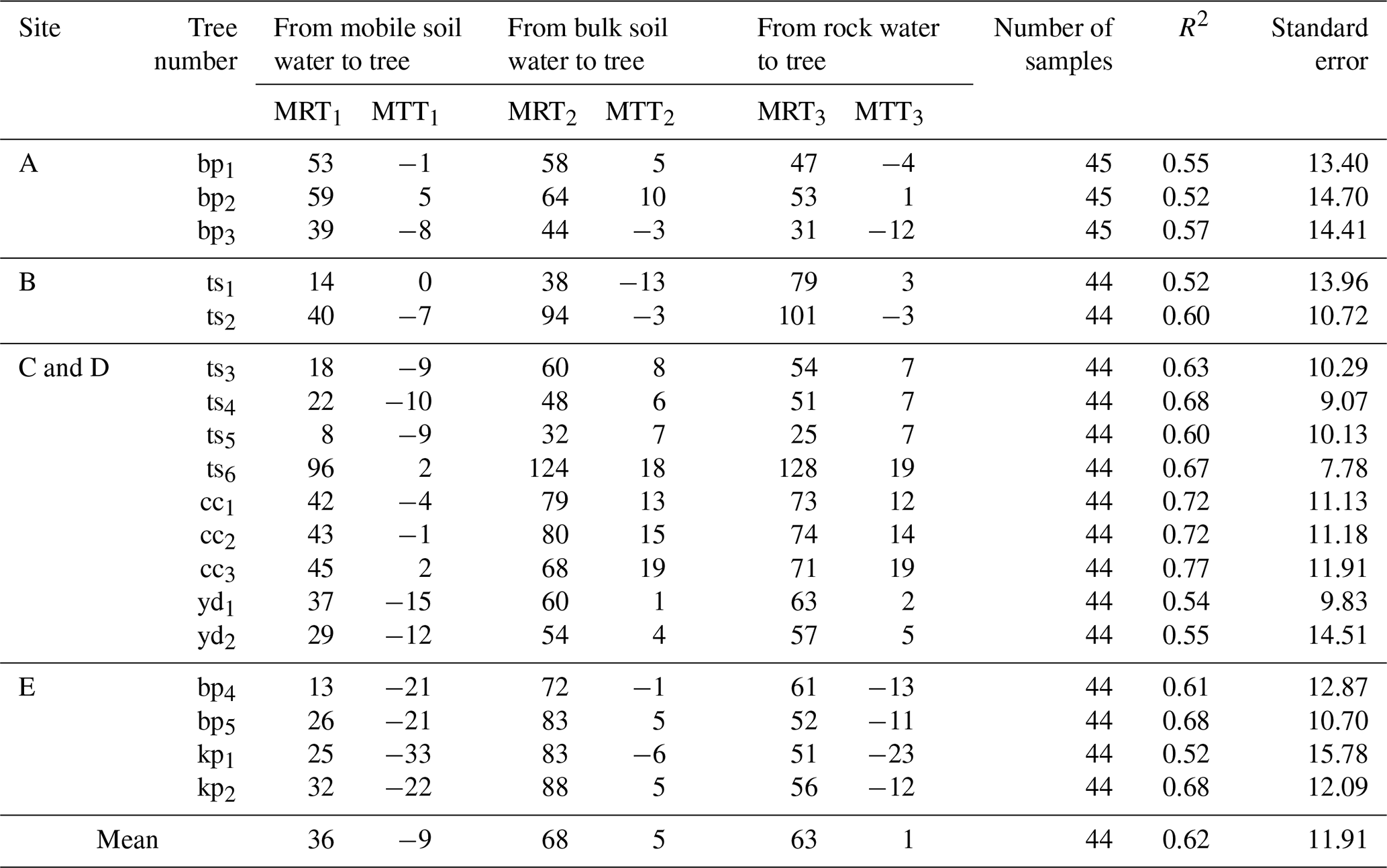
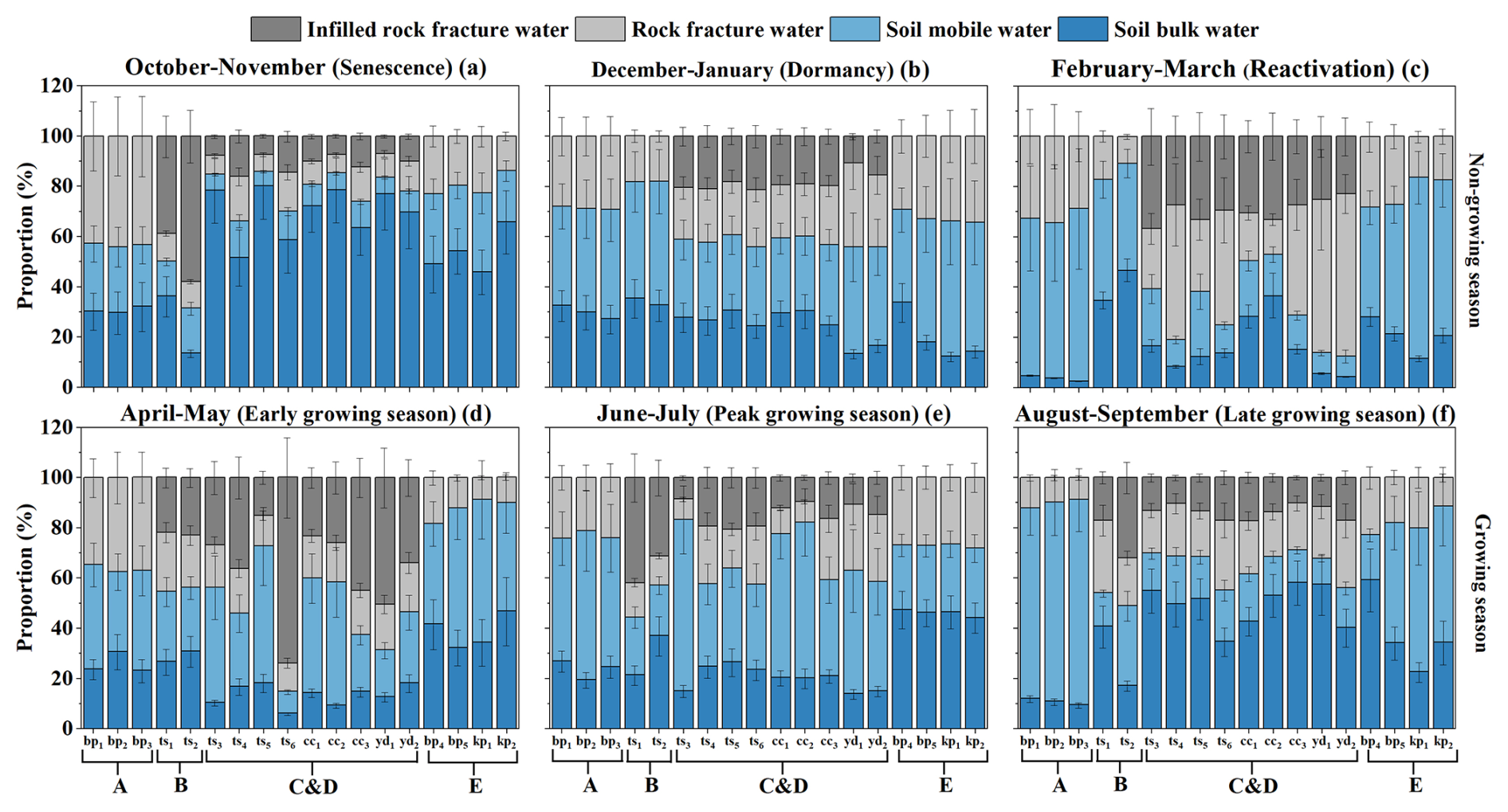
Figure D1Seasonal variation in the proportional contributions of different water sources to individual tree xylem water across sites. Error bars indicate ±1 standard deviation (SD) derived from the MixSIAR model outputs.
To validate the results of this study and ensure reproducibility, the datasets are accessible on Zenodo (https://doi.org/10.5281/zenodo.14827769, https://doi.org/10.5281/zenodo.16810833) and are publicly available for download (Liu, 2025a, b). The underlying research codes used in this study are publicly available on Zenodo at https://doi.org/10.5281/zenodo.14565556 (Liu, 2024).
XL, XC, and JJM conceived the study. XL, WL, and ZZ curated the data. XL and JJM performed the formal analysis. XL and WL carried out the investigation. XL, XC, and JJM developed the methodology. XC, TP, and JJM provided resources. XL developed the software and performed the validation. XL and JJM prepared the original draft. XL, XC, ZZ, and JJM reviewed and edited the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank Kim Janzen for conducting all isotopic analyses, and Cody Millar and Hongxiu Wang for their assistance with sample extractions at the University of Saskatchewan. We also gratefully acknowledge Bingcheng Si, Qin Liu and Ha Fu for their valuable comments on the manuscript. The Department of Soil Science at the University of Saskatchewan is thanked for hosting the senior author while the paper was prepared. We thank the Puding Karst Ecosystem Research Station, Chinese Academy of Sciences, for providing field experimental support, and Dr. Jia Chen for kindly sharing the meteorological data.
This research was funded by the National Natural Science Foundation of China (grant nos. 42030506, 42261144672) and the China Scholarship Council (grant no. 202306250097).
This paper was edited by Hongkai Gao and reviewed by two anonymous referees.
Allen, S. T., Kirchner, J. W., and Goldsmith, G. R.: Predicting Spatial Patterns in Precipitation Isotope (δ2H and δ18O) Seasonality Using Sinusoidal Isoscapes, Geophys. Res. Lett., 45, 4859–4868, https://doi.org/10.1029/2018GL077458, 2018.
Asadollahi, M., Stumpp, C., Rinaldo, A., and Benettin, P.: Transport and Water Age Dynamics in Soils: A Comparative Study of Spatially Integrated and Spatially Explicit Models, Water Resour. Res., 56, e2019WR025539, https://doi.org/10.1029/2019WR025539, 2020.
Barbeta, A., Mejía-Chang, M., Ogaya, R., Voltas, J., Dawson, T. E., and Peñuelas, J.: The combined effects of a long-term experimental drought and an extreme drought on the use of plant-water sources in a Mediterranean forest, Glob. Change Biol., 21, 1213–1225, https://doi.org/10.1111/gcb.12785, 2015.
Benettin, P., Soulsby, C., Birkel, C., Tetzlaff, D., Botter, G., and Rinaldo, A.: Using SAS functions and high-resolution isotope data to unravel travel time distributions in headwater catchments, Water Resour. Res., 53, 1864–1878, https://doi.org/10.1002/2016WR020117, 2017.
Bond, B. J., Meinzer, F. C., and Brooks, J. R.: How Trees Influence the Hydrological Cycle in Forest Ecosystems, in: Hydroecology and Ecohydrology, 7–35, https://doi.org/10.1002/9780470010198.ch2, 2008.
Brooks, J. R., Barnard, H. R., Coulombe, R., and McDonnell, J. J.: Ecohydrologic separation of water between trees and streams in a Mediterranean climate, Nat. Geosci., 3, 100–104, https://doi.org/10.1038/ngeo722, 2010.
Cai, L., Xiong, K., Liu, Z., Li, Y., and Fan, B.: Seasonal variations of plant water use in the karst desertification control, Sci. Total Environ., 885, 163778, https://doi.org/10.1016/j.scitotenv.2023.163778, 2023.
Cai, L., Xiong, K., Liu, Z., Li, Y., Zhu, D., Mu, Y., Chen, Y., Zhou, R., and Yan, X.: Water competition among coexisting plants in rock-dominated habitats of subtropical karst desertification, Agr. Ecosyst. Environ., 381, 109419, https://doi.org/10.1016/j.agee.2024.109419, 2025.
Carrière, S., Martin-StPaul, N., Cakpo, C., Patris, N., Gillon, M., Chalikakis, K., Doussan, C., Olioso, A., Babic, M., Jouineau, A., Simioni, G., and Davi, H.: The role of deep vadose zone water in tree transpiration during drought periods in karst settings – Insights from isotopic tracing and leaf water potential, Sci. Total Environ., 699, 134332, https://doi.org/10.1016/j.scitotenv.2019.134332, 2019.
Chen, H. and Wang, S.: Accelerated Transition Between Dry and Wet Periods in a Warming Climate, Geophys. Res. Lett., 49, e2022GL099766, https://doi.org/10.1029/2022GL099766, 2022.
Chen, H., Liu, J., Wang, K., and Zhang, W.: Spatial distribution of rock fragments on steep hillslopes in karst region of northwest Guangxi, China, CATENA, 84, 21–28, https://doi.org/10.1016/j.catena.2010.08.012, 2011.
Chen, L., Brun, P., Buri, P., Fatichi, S., Gessler, A., McCarthy, M., Pellicciotti, F., Stocker, B., and Karger, D. N.: Global increase in the occurrence and impact of multiyear droughts, Science, 387, 278–284, https://doi.org/10.1126/science.ado4245, 2025.
Compilation Committee of the Assessment Report on Climate Change in Southwest China: Summary for policymakers and executive summary of assessment report on climate change in Southwest China 2020, China Meteorological Press, Beijing, China, 2021 (in Chinese).
Darrouzet-Nardi, A., D'Antonio, C. M., and Dawson, T. E.: Depth of water acquisition by invading shrubs and resident herbs in a Sierra Nevada meadow, Plant Soil, 285, 31–43, https://doi.org/10.1007/s11104-005-4453-z, 2006.
Deng, Y., Kuo, Y.-M., Jiang, Z., Qin, X., and Jin, Z.: Using stable isotopes to quantify water uptake by Cyclobalanopsis glauca in typical clusters of karst peaks in China, Environ. Earth Sci., 74, 1039–1046, https://doi.org/10.1007/s12665-014-3780-x, 2015.
Deng, Y., Ke, J., Wu, S., Jiang, G., Jiang, Z., and Zhu, A.: Responses of plant water uptake to groundwater depth in limestone outcrops, J. Hydrol., 590, 125377, https://doi.org/10.1016/j.jhydrol.2020.125377, 2020.
Deng, Z., Guan, H., Hutson, J., Forster, M. A., Wang, Y., and Simmons, C. T.: A vegetation-focused soil-plant-atmospheric continuum model to study hydrodynamic soil-plant water relations, Water Resour. Res., 53, 4965–4983, https://doi.org/10.1002/2017WR020467, 2017.
Ding, Y., Nie, Y., Chen, H., Wang, K., and Querejeta, J. I.: Water uptake depth is coordinated with leaf water potential, water-use efficiency and drought vulnerability in karst vegetation, New Phytol., 229, 1339–1353, https://doi.org/10.1111/nph.16971, 2021.
Ding, Y., Nie, Y., Zhang, W., Wang, K., Wen, L., and Chen, H.: Unveiling plant adaptations: How limestone outperforms dolomite in water supply capacity, Journal of Hydrology: Regional Studies, 61, 102641, https://doi.org/10.1016/j.ejrh.2025.102641, 2025.
Epstein, S. and Mayeda, T.: Variation of O18 content of waters from natural sources, Geochim. Cosmochim. Ac., 4, 213–224, https://doi.org/10.1016/0016-7037(53)90051-9, 1953.
Estrada-Medina, H., Graham, R. C., Allen, M. F., Jiménez-Osornio, J. J., and Robles-Casolco, S.: The importance of limestone bedrock and dissolution karst features on tree root distribution in northern Yucatán, México, Plant Soil, 362, 37–50, https://doi.org/10.1007/s11104-012-1175-x, 2013.
Evaristo, J., Jasechko, S., and McDonnell, J. J.: Global separation of plant transpiration from groundwater and streamflow, Nature, 525, 91–94, https://doi.org/10.1038/nature14983, 2015.
Evaristo, J., McDonnell, J. J., and Clemens, J.: Plant source water apportionment using stable isotopes: A comparison of simple linear, two-mixing model approaches, Hydrol. Process., 31, 3750–3758, https://doi.org/10.1002/hyp.11233, 2017.
Fabiani, G., Klaus, J., and Penna, D.: The influence of hillslope topography on beech water use: a comparative study in two different climates, Hydrol. Earth Syst. Sci., 28, 2683–2703, https://doi.org/10.5194/hess-28-2683-2024, 2024.
Fan, B., Xiong, K., and Liu, Z.: Forest Plant Water Utilization and the Eco-Hydrological Regulation in the Karst Desertification Control Drainage Area, Forests, 14, 747, https://doi.org/10.3390/f14040747, 2023.
Fan, Y. and Miguez-Macho, G.: Infiltration depth, rooting depth, and regolith flushing-A global perspective, PNAS Nexus, 3, pgae514, https://doi.org/10.1093/pnasnexus/pgae514, 2024.
Finkenbiner, C. E., Good, S., Brooks, J., Allen, S., and Sasidharan, S.: The physical basis for ecohydrologic separation: the roles of soil hydraulics and climate, https://doi.org/10.21203/rs.3.rs-1024190/v1, 2021.
Finkenbiner, C. E., Good, S. P., Renée Brooks, J., Allen, S. T., and Sasidharan, S.: The extent to which soil hydraulics can explain ecohydrological separation, Nat. Commun., 13, 6492, https://doi.org/10.1038/s41467-022-34215-7, 2022.
Fu, H., Neil, E. J., Liu, J., and Si, B.: A Continuous Root Water Uptake Isotope Mixing Model, Water Resour. Res., 60, e2023WR036852, https://doi.org/10.1029/2023WR036852, 2024.
Gai, H., Shi, P., and Li, Z.: Untangling the Uncertainties in Plant Water Source Partitioning With Isotopes, Water Resour. Res., 59, e2022WR033849, https://doi.org/10.1029/2022WR033849, 2023.
Geris, J., Tetzlaff, D., McDonnell, J., Anderson, J., Paton, G., and Soulsby, C.: Ecohydrological separation in wet, low energy northern environments? A preliminary assessment using different soil water extraction techniques, Hydrol. Process., 29, 5139–5152, https://doi.org/10.1002/hyp.10603, 2015.
Hahm, W. J., Rempe, D. M., Dralle, D. N., Dawson, T. E., and Dietrich, W. E.: Oak Transpiration Drawn From the Weathered Bedrock Vadose Zone in the Summer Dry Season, Water Resour. Res., 56, e2020WR027419, https://doi.org/10.1029/2020WR027419, 2020.
Hahm, W. J., Dralle, D. N., Sanders, M., Bryk, A. B., Fauria, K. E., Huang, M. H., Hudson-Rasmussen, B., Nelson, M. D., Pedrazas, M. A., Schmidt, L., Whiting, J., Dietrich, W. E., and Rempe, D. M.: Bedrock Vadose Zone Storage Dynamics Under Extreme Drought: Consequences for Plant Water Availability, Recharge, and Runoff, Water Resour. Res., 58, e2021WR031781, https://doi.org/10.1029/2021WR031781, 2022.
Hartmann, A., Goldscheider, N., Wagener, T., Lange, J., and Weiler, M.: Karst water resources in a changing world: Review of hydrological modeling approaches, Rev. Geophys., 52, 218–242, https://doi.org/10.1002/2013RG000443, 2014.
Hasenmueller, E. A., Gu, X., Weitzman, J. N., Adams, T. S., Stinchcomb, G. E., Eissenstat, D. M., Drohan, P. J., Brantley, S. L., and Kaye, J. P.: Weathering of rock to regolith: The activity of deep roots in bedrock fractures, Geoderma, 300, 11–31, https://doi.org/10.1016/j.geoderma.2017.03.020, 2017.
He, K., Chen, X., Zhou, J., Zhao, D., and Yu, X.: Compound successive dry-hot and wet extremes in China with global warming and urbanization, J. Hydrol., 636, 131332, https://doi.org/10.1016/j.jhydrol.2024.131332, 2024.
Hervé-Fernández, P., Oyarzún, C., Brumbt, C., Huygens, D., Bodé, S., Verhoest, N. E. C., and Boeckx, P.: Assessing the “two water worlds” hypothesis and water sources for native and exotic evergreen species in south-central Chile, Hydrol. Process., 30, 4227–4241, https://doi.org/10.1002/hyp.10984, 2016.
Howarth, F. and Bishop, B.: Why the delay in recognizing terrestrial obligate cave species in the tropics?, Int. J. Speleol., 52, https://doi.org/10.5038/1827-806X.52.1.2446, 2023.
Hubbert, K., Beyers, J., and Graham, R.: Roles of weathered bedrock and soil in seasonal water relations of Pinus Jeffreyi and Arctostaphylos patula, Can. J. Forest Res., 31, 1947–1957, https://doi.org/10.1139/x01-136, 2011.
Jiang, Z., Liu, H., Wang, H., Peng, J., Meersmans, J., Green, S. M., Quine, T. A., Wu, X., and Song, Z.: Bedrock geochemistry influences vegetation growth by regulating the regolith water holding capacity, Nat. Commun., 11, 2392, https://doi.org/10.1038/s41467-020-16156-1, 2020.
Jiménez-Rodríguez, C. D., Sulis, M., and Schymanski, S.: Exploring the role of bedrock representation on plant transpiration response during dry periods at four forested sites in Europe, Biogeosciences, 19, 3395–3423, https://doi.org/10.5194/bg-19-3395-2022, 2022.
Kirchner, J. W.: Aggregation in environmental systems – Part 1: Seasonal tracer cycles quantify young water fractions, but not mean transit times, in spatially heterogeneous catchments, Hydrol. Earth Syst. Sci., 20, 279–297, https://doi.org/10.5194/hess-20-279-2016, 2016.
Koeniger, P., Marshall, J. D., Link, T., and Mulch, A.: An inexpensive, fast, and reliable method for vacuum extraction of soil and plant water for stable isotope analyses by mass spectrometry, Rapid Commun. Mass Sp., 25, 3041–3048, https://doi.org/10.1002/rcm.5198, 2011.
Korboulewsky, N., Tétégan, M., Samouelian, A., and Cousin, I.: Plants use water in the pores of rock fragments during drought, Plant Soil, 454, 35–47, https://doi.org/10.1007/s11104-020-04425-3, 2020.
Landwehr, J. M. and Coplen, T. B.: Line-conditioned excess: a new method for characterizing stable hydrogen and oxygen isotope ratios in hydrologic systems (IAEA-CN-118/56), in: Isotopes in Environmental Studies, IAEA, Vienna, 132–135, 2006.
Leite, P. A. M., Rempe, D. M., McInnes, K. J., Schmidt, L. M., Walker, J. W., Olariu, H. G., and Wilcox, B. P.: Trees Enhance Rock Moisture Storage: A Major Pool in Karst Drylands and Crucial During Droughts, Water Resour. Res., 61, e2024WR038692, https://doi.org/10.1029/2024WR038692, 2025.
Li, Y., Wang, S., Peng, T., Zhao, G., and Dai, B.: Hydrological characteristics and available water storage of typical karst soil in SW China under different soil–rock structures, Geoderma, 438, 116633, https://doi.org/10.1016/j.geoderma.2023.116633, 2023.
Lin, W., Wen, C., Wen, Z., and Gang, H.: Drought in Southwest China: A Review, Atmospheric and Oceanic Science Letters, 8, 339–344, https://doi.org/10.3878/AOSL20150043, 2015.
Liu, Q., Wang, T., Liu, C.-q., Mikouendanandi, E. M. R. B., Chen, X., Peng, T., and Zhang, L.: Characterizing the spatiotemporal dynamics of shallow soil water stable isotopic compositions on a karst hillslope in Southwestern China, J. Hydrol., 610, 127964, https://doi.org/10.1016/j.jhydrol.2022.127964, 2022.
Liu, W., M. Behzad, H., Luo, Z., Huang, L., and Nie, Y.-P.: Species-specific leaf iso/anisohydric tendencies and root distribution shape transpiration patterns across karst habitats, https://doi.org/10.22541/au.171474150.06111999/v1, 2024a.
Liu, W., Nie, Y., Behzad, H. M., Luo, Z., Huang, L., Wang, Z., Huang, Y., and Chen, H.: Trade-offs between root water uptake and xylem-leaf water-use traits underpin contrasting plant adaptation strategies in limestone and dolomite habitats, Tree Physiol., 46, https://doi.org/10.1093/treephys/tpag013, 2026.
Liu, X.: Chronic nitrogen legacy in the aquifers of China, Zenodo [code], https://doi.org/10.5281/zenodo.14565556, 2024.
Liu, X.: Guizhou Karst Rock Fracture, Soil Moisture, Sap Flow, and Meteorological Data, Zenodo [data set], https://doi.org/10.5281/zenodo.14827769, 2025a.
Liu, X.: Stable isotope (δ18O, δD) dataset of soil water, rock fracture water, precipitation, and xylem water in a subtropical karst ecosystem, southwest China (2022–2025), Zenodo [data set], https://doi.org/10.5281/zenodo.16810833, 2025b.
Liu, X., Yue, F.-J., Wong, W. W., Guo, T.-L., and Li, S.-L.: Unravelling nitrate transformation mechanisms in karst catchments through the coupling of high-frequency sensor data and machine learning, Water Res., 267, 122507, https://doi.org/10.1016/j.watres.2024.122507, 2024b.
Liu, X., Liu, W., Chen, X., Wang, L., Zhang, Z., and Peng, T.: Estimating fracture characteristics and hydraulic conductivity from slug tests in epikarst of southwest China, Journal of Hydrology: Regional Studies, 53, 101777, https://doi.org/10.1016/j.ejrh.2024.101777, 2024c.
Liu, X., Chen, X., Zhang, Z., Liu, W., Gao, F., Cheng, Q., Chen, J., and Peng, T.: The Role of Rock Fractures as a Water Source for Trees Growing in Karst, Water Resour. Res., 61, e2024WR039588, https://doi.org/10.1029/2024WR039588, 2025.
Liu, Y., Fang, Y., Hu, H., Tian, F., Dong, Z., and Khan, M. Y. A.: Ecohydrological Separation Hypothesis: Review and Prospect, Water, 12, 2077, https://doi.org/10.3390/w12082077, 2020.
Liu, Z., Liu, Z., Jia, G., Liu, Z., and Yu, X.: Employing stable isotopes to reveal temporal trajectories of water travelling through the soil–plant-atmosphere continuum, J. Hydrol., 644, 132058, https://doi.org/10.1016/j.jhydrol.2024.132058, 2024d.
Luo, J., Luo, W., Liu, J., Wang, Y., Li, Z., Tao, J., and Liu, J.: Karst fissures mitigate the negative effects of drought on plant growth and photosynthetic physiology, Oecologia, 205, 69–80, https://doi.org/10.1007/s00442-024-05556-5, 2024a.
Luo, Z., Guan, H., Zhang, X., Xu, X., Dai, J., and Hua, M.: Examination of the ecohydrological separation hypothesis in a humid subtropical area: Comparison of three methods, J. Hydrol., 571, 642–650, https://doi.org/10.1016/j.jhydrol.2019.02.019, 2019.
Luo, Z., Nie, Y., Chen, H., Guan, H., Zhang, X., and Wang, K.: Water Age Dynamics in Plant Transpiration: The Effects of Climate Patterns and Rooting Depth, Water Resour. Res., 59, e2022WR033566, https://doi.org/10.1029/2022WR033566, 2023.
Luo, Z., Fan, J., Shao, M., Yang, Q., and Li, M.: Rock moisture reinforces belowground water storage under different precipitation scenarios and vegetation coverage, J. Hydrol., 636, 131276, https://doi.org/10.1016/j.jhydrol.2024.131276, 2024b.
Małoszewski, P. and Zuber, A.: Determining the turnover time of groundwater systems with the aid of environmental tracers: 1. Models and their applicability, J. Hydrol., 57, 207–231, https://doi.org/10.1016/0022-1694(82)90147-0, 1982.
McCole, A. A. and Stern, L. A.: Seasonal water use patterns of Juniperus ashei on the Edwards Plateau, Texas, based on stable isotopes in water, J. Hydrol., 342, 238–248, https://doi.org/10.1016/j.jhydrol.2007.05.024, 2007.
McCormick, E. L., Dralle, D. N., Hahm, W. J., Tune, A. K., Schmidt, L. M., Chadwick, K. D., and Rempe, D. M.: Widespread woody plant use of water stored in bedrock, Nature, 597, 225–229, https://doi.org/10.1038/s41586-021-03761-3, 2021.
McDonnell, J. J.: The two water worlds hypothesis: ecohydrological separation of water between streams and trees?, WIREs Water, 1, 323–329, https://doi.org/10.1002/wat2.1027, 2014.
McGuire, K. J., DeWalle, D. R., and Gburek, W. J.: Evaluation of mean residence time in subsurface waters using oxygen-18 fluctuations during drought conditions in the mid-Appalachians, J. Hydrol., 261, 132–149, https://doi.org/10.1016/S0022-1694(02)00006-9, 2002.
Midwood, A. J., Boutton, T. W., Archer, S. R., and Watts, S. E.: Water use by woody plants on contrasting soils in a savanna parkland: assessment with δ2H and δ18O, Plant Soil, 205, 13–24, https://doi.org/10.1023/A:1004355423241, 1998.
Moore, J. W. and Semmens, B. X.: Incorporating uncertainty and prior information into stable isotope mixing models, Ecol. Lett., 11, 470–480, https://doi.org/10.1111/j.1461-0248.2008.01163.x, 2008.
Morrison, J., Brockwell, T., Merren, T., Fourel, F., and Phillips, A. M.: On-line high-precision stable hydrogen isotopic analyses on nanoliter water samples, Anal. Chem., 73, 3570–3575, https://doi.org/10.1021/ac001447t, 2001.
Nardini, A., Petruzzellis, F., Marusig, D., Tomasella, M., Natale, S., Altobelli, A., Calligaris, C., Floriddia, G., Cucchi, F., Forte, E., and Zini, L.: Water “on the rocks”: a summer drink for thirsty trees?, New Phytol., 229, 199–212, https://doi.org/10.1111/nph.16859, 2021.
Nardini, A., Tomasella, M., and Di Bert, S.: Bedrock: the hidden water reservoir for trees challenged by drought, Trees, 38, 1–11, https://doi.org/10.1007/s00468-023-02482-6, 2024.
Nie, Y.-P., Chen, H.-S., Wang, K.-L., and Yang, J.: Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China, J. Hydrol., 420, 264–274, https://doi.org/10.1016/j.jhydrol.2011.12.011, 2012.
Ning, J., Liu, X., Wu, X., Yang, H., Ma, J., and Cao, J.: The Effect of Bedrock Differences on Plant Water Use Strategies in Typical Karst Areas of Southwest China, Land, 12, https://doi.org/10.3390/land12010012, 2023.
Oshun, J., Dietrich, W. E., Dawson, T. E., and Fung, I.: Dynamic, structured heterogeneity of water isotopes inside hillslopes, Water Resour. Res., 52, 164–189, https://doi.org/10.1002/2015WR017485, 2016.
Parnell, A. C., Phillips, D. L., Bearhop, S., Semmens, B. X., Ward, E. J., Moore, J. W., Jackson, A. L., Grey, J., Kelly, D. J., and Inger, R.: Bayesian stable isotope mixing models, Environmetrics, 24, 387–399, https://doi.org/10.1002/env.2221, 2013.
Parry, J. T.: The limestone pavements of northwestern England, Canadian Geographies, 4, 14–21, https://doi.org/10.1111/j.1541-0064.1960.tb01836.x, 1960.
Pawlik, Ł., Phillips, J. D., and amonil, P.: Roots, rock, and regolith: Biomechanical and biochemical weathering by trees and its impact on hillslopes – A critical literature review, Earth-Sci. Rev., 159, 142–159, https://doi.org/10.1016/j.earscirev.2016.06.002, 2016.
Peng, X., Wang, X., Dai, Q., Ding, G., and Li, C.: Soil structure and nutrient contents in underground fissures in a rock-mantled slope in the karst rocky desertification area, Environ. Earth Sci., 79, https://doi.org/10.1007/s12665-019-8708-z, 2019.
Peng, X., Dai, Q., Ding, G., Shi, D., and Li, C.: Impact of vegetation restoration on soil properties in near-surface fissures located in karst rocky desertification regions, Soil Till. Res., 200, 104620, https://doi.org/10.1016/j.still.2020.104620, 2020.
Philip, J. R.: Plant Water Relations: Some Physical Aspects, Annu. Rev. Plant Phys., 17, 245–268, 1966.
Preisler, Y., Tatarinov, F., Grünzweig, J. M., Bert, D., Ogée, J., Wingate, L., Rotenberg, E., Rohatyn, S., Her, N., Moshe, I., Klein, T., and Yakir, D.: Mortality versus survival in drought-affected Aleppo pine forest depends on the extent of rock cover and soil stoniness, Funct. Ecol., 33, 901–912, https://doi.org/10.1111/1365-2435.13302, 2019.
Putman, A. L. and Bowen, G. J.: Technical Note: A global database of the stable isotopic ratios of meteoric and terrestrial waters, Hydrol. Earth Syst. Sci., 23, 4389–4396, https://doi.org/10.5194/hess-23-4389-2019, 2019.
Querejeta, J. I., Estrada-Medina, H., Allen, M. F., Jiménez-Osornio, J. J., and Ruenes, R.: Utilization of bedrock water by Brosimum alicastrum trees growing on shallow soil atop limestone in a dry tropical climate, Plant Soil, 287, 187–197, https://doi.org/10.1007/s11104-006-9065-8, 2006.
Reddy, M. M., Schuster, P., Kendall, C., and Reddy, M. B.: Characterization of surface and ground water δ18O seasonal variation and its use for estimating groundwater residence times, Hydrol. Process., 20, 1753–1772, https://doi.org/10.1002/hyp.5953, 2006.
Rempe, D. M. and Dietrich, W. E.: Direct observations of rock moisture, a hidden component of the hydrologic cycle, P. Natl. Acad. Sci. USA, 115, 2664–2669, https://doi.org/10.1073/pnas.1800141115, 2018.
Rong, L., Chen, X., Chen, X., Wang, S., and Du, X.: Isotopic analysis of water sources of mountainous plant uptake in a karst plateau of southwest China, Hydrol. Process., 25, 3666–3675, https://doi.org/10.1002/hyp.8093, 2011.
Salomón, R. L., Peters, R. L., Zweifel, R., Sass-Klaassen, U. G. W., Stegehuis, A. I., Smiljanic, M., Poyatos, R., Babst, F., Cienciala, E., Fonti, P., Lerink, B. J. W., Lindner, M., Martinez-Vilalta, J., Mencuccini, M., Nabuurs, G.-J., van der Maaten, E., von Arx, G., Bär, A., Akhmetzyanov, L., Balanzategui, D., Bellan, M., Bendix, J., Berveiller, D., Blaenec, M., Čada, V., Carraro, V., Cecchini, S., Chan, T., Conedera, M., Delpierre, N., Delzon, S., Ditmarová, Ĺ, Dolezal, J., Dufrêne, E., Edvardsson, J., Ehekircher, S., Forner, A., Frouz, J., Ganthaler, A., Gryc, V., Güney, A., Heinrich, I., Hentschel, R., Janda, P., Ježík, M., Kahle, H.-P., Knüsel, S., Krejza, J., Kuberski, Ł., Kučera, J., Lebourgeois, F., Mikolá, M., Matula, R., Mayr, S., Oberhuber, W., Obojes, N., Osborne, B., Paljakka, T., Plichta, R., Rabbel, I., Rathgeber, C. B. K., Salmon, Y., Saunders, M., Scharnweber, T., Sitková, Z., Stangler, D. F., Stereńczak, K., Stojanović, M., Střelcová, K., Světlík, J., Svoboda, M., Tobin, B., Trotsiuk, V., Urban, J., Valladares, F., Vavrčík, H., Vejpustková, M., Walthert, L., Wilmking, M., Zin, E., Zou, J., and Steppe, K.: The 2018 European heatwave led to stem dehydration but not to consistent growth reductions in forests, Nat. Commun., 13, 28, https://doi.org/10.1038/s41467-021-27579-9, 2022.
Salve, R., Rempe, D. M., and Dietrich, W. E.: Rain, rock moisture dynamics, and the rapid response of perched groundwater in weathered, fractured argillite underlying a steep hillslope, Water Resour. Res., 48, https://doi.org/10.1029/2012WR012583, 2012.
Schoeman, J. L., Kruger, M. M., and Loock, A. H.: Water-holding capacity of rock fragments in rehabilitated opencast mine soils, South African Journal of Plant Soil, 14, 98–102, https://doi.org/10.1080/02571862.1997.10635089, 1997.
Schwinning, S.: The ecohydrology of roots in rocks, Ecohydrology, 3, 238–245, https://doi.org/10.1002/eco.134, 2010.
Schwinning, S.: A critical question for the critical zone: how do plants use rock water?, Plant Soil, 454, 49–56, https://doi.org/10.1007/s11104-020-04648-4, 2020.
Snelgrove, J. R., Buttle, J. M., Kohn, M. J., and Tetzlaff, D.: Co-evolution of xylem water and soil water stable isotopic composition in a northern mixed forest biome, Hydrol. Earth Syst. Sci., 25, 2169–2186, https://doi.org/10.5194/hess-25-2169-2021, 2021.
Sprenger, M. and Allen, S. T.: What Ecohydrologic Separation Is and Where We Can Go With It, Water Resour. Res., 56, e2020WR027238, https://doi.org/10.1029/2020WR027238, 2020.
Sprenger, M., Leistert, H., Gimbel, K., and Weiler, M.: Illuminating hydrological processes at the soil-vegetation-atmosphere interface with water stable isotopes, Rev. Geophys., 54, 674–704, https://doi.org/10.1002/2015RG000515, 2016.
Sprenger, M., Tetzlaff, D., Buttle, J., Laudon, H., Leistert, H., Mitchell, C. P. J., Snelgrove, J., Weiler, M., and Soulsby, C.: Measuring and Modeling Stable Isotopes of Mobile and Bulk Soil Water, Vadose Zone J., 17, 170149, https://doi.org/10.2136/vzj2017.08.0149, 2018.
Sprenger, M., Llorens, P., Cayuela, C., Gallart, F., and Latron, J.: Mechanisms of consistently disjunct soil water pools over (pore) space and time, Hydrol. Earth Syst. Sci., 23, 2751–2762, https://doi.org/10.5194/hess-23-2751-2019, 2019.
Stewart, M. K. and McDonnell, J. J.: Modeling Base Flow Soil Water Residence Times From Deuterium Concentrations, Water Resour. Res., 27, 2681–2693, https://doi.org/10.1029/91WR01569, 1991.
Stock, B. C. and Semmens, B. X.: Unifying error structures in commonly used biotracer mixing models, Ecology, 97, 2562–2569, https://doi.org/10.1002/ecy.1517, 2016.
Tang, K. and Feng, X.: The effect of soil hydrology on the oxygen and hydrogen isotopic compositions of plants' source water, Earth Planet. Sc. Lett., 185, 355–367, https://doi.org/10.1016/S0012-821X(00)00385-X, 2001.
Tong, X., Brandt, M., Yue, Y., Horion, S., Wang, K., Keersmaecker, W. D., Tian, F., Schurgers, G., Xiao, X., Luo, Y., Chen, C., Myneni, R., Shi, Z., Chen, H., and Fensholt, R.: Increased vegetation growth and carbon stock in China karst via ecological engineering, Nature Sustainability, 1, 44–50, https://doi.org/10.1038/s41893-017-0004-x, 2018.
Vargas, A. I., Schaffer, B., Yuhong, L., and Sternberg, L. d. S. L.: Testing plant use of mobile vs. immobile soil water sources using stable isotope experiments, New Phytol., 215, 582–594, https://doi.org/10.1111/nph.14616, 2017.
Vrettas, M. D. and Fung, I. Y.: Sensitivity of transpiration to subsurface properties: Exploration with a 1-D model, J. Adv. Model. Earth Sy., 9, 1030–1045, https://doi.org/10.1002/2016MS000901, 2017.
Wang, H., Yu, H., He, D., Li, M., Si, B., McDonnell, J. J., and Nehemy, M. F.: Cryogenic vacuum distillation vs. Cavitron methods in ecohydrology: Extraction protocol effects on plant water isotopic values, J. Hydrol., 642, 131853, https://doi.org/10.1016/j.jhydrol.2024.131853, 2024a.
Wang, J., Lu, N., and Fu, B.: Inter-comparison of stable isotope mixing models for determining plant water source partitioning, Sci. Total Environ., 666, 685–693, https://doi.org/10.1016/j.scitotenv.2019.02.262, 2019a.
Wang, K., Zhang, C., Chen, H., Yue, Y., Zhang, W., Zhang, M., Qi, X., and Fu, Z.: Karst landscapes of China: patterns, ecosystem processes and services, Landscape Ecol., 34, 2743–2763, https://doi.org/10.1007/s10980-019-00912-w, 2019b.
Wang, K., Liu, Z., Zeng, T., Wang, F., Shen, W., and Shao, J.: Performance of enhanced geothermal system with varying injection-production parameters and reservoir properties, Appl. Therm. Eng., 207, 118160, https://doi.org/10.1016/j.applthermaleng.2022.118160, 2022.
Wang, L., Cardenas, M. B., Slottke, D. T., Ketcham, R. A., and Sharp Jr., J. M.: Modification of the Local Cubic Law of fracture flow for weak inertia, tortuosity, and roughness, Water Resour. Res., 51, https://doi.org/10.1002/2014WR015815, 2015.
Wang, P., Song, X., Han, D., Zhang, Y., and Liu, X.: A study of root water uptake of crops indicated by hydrogen and oxygen stable isotopes: A case in Shanxi Province, China, Agr. Water Manage., 97, 475–482, https://doi.org/10.1016/j.agwat.2009.11.008, 2010.
Wang, S., Yan, Y., Zhao, Y., Fu, Z., and Chen, H.: Co-evolution among soil thickness, epikarst weathering degree, and runoff characteristics on a subtropical karst hillslope, J. Hydrol., 628, 130499, https://doi.org/10.1016/j.jhydrol.2023.130499, 2024b.
Wei, S., Chu, X., Sun, B., Yuan, W., Song, W., Zhao, M., Wang, X., Li, P., and Han, G.: Climate warming negatively affects plant water-use efficiency in a seasonal hydroperiod wetland, Water Res., 242, 120246, https://doi.org/10.1016/j.watres.2023.120246, 2023.
Williams, P. W.: The role of the epikarst in karst and cave hydrogeology: a review, Int. J. Speleol., 37, 1–10, https://doi.org/10.5038/1827-806X.37.1.1, 2008.
Wolfsberg, A.: Rock Fractures and Fluid Flow: Contemporary Understanding and Applications, Eos T. Am. Geophys. Un., 78, 569–573, https://doi.org/10.1029/97EO00345, 1997.
Wu, H., Fan, H., Li, J., Yue, F.-J., Lian, E., Fu, C., Lei, R., Ding, M., Liu, J., and Li, X.-Y.: Reproducing surface water isoscapes of δ18O and δ2H across China: A machine learning approach, J. Hydrol., 638, 131565, https://doi.org/10.1016/j.jhydrol.2024.131565, 2024a.
Wu, Z., Behzad, H. M., He, Q., Wu, C., Bai, Y., and Jiang, Y.: Seasonal transpiration dynamics of evergreen Ligustrum lucidum linked with water source and water-use strategy in a limestone karst area, southwest China, J. Hydrol., 597, 126199, https://doi.org/10.1016/j.jhydrol.2021.126199, 2021.
Wu, Z., Lv, T., Zeng, S., Zhang, X., Wu, H., Luo, S., Bai, Y., Zhang, Y., and Jiang, Y.: Differential responses in water-use strategies of evergreen (Ligustrum lucidum) and deciduous (Robinia pseudoacacia) trees to tunnel excavation in a subtropical karst trough valley, J. Hydrol., 636, 131323, https://doi.org/10.1016/j.jhydrol.2024.131323, 2024b.
Xu, W., Xia, X., Piao, S., Wu, D., Li, W., Yang, S., and Yuan, W.: Weakened Increase in Global Near-Surface Water Vapor Pressure During the Last 20 Years, Geophys. Res. Lett., 51, e2023GL107909, https://doi.org/10.1029/2023GL107909, 2024.
Xu, X., Yan, Y., Dai, Q., Yi, X., Hu, Z., and Cen, L.: Spatial and temporal dynamics of rainfall erosivity in the karst region of southwest China: Interannual and seasonal changes, CATENA, 221, 106763, https://doi.org/10.1016/j.catena.2022.106763, 2023.
Xu, X., Zhao, Z., and Skrzypek, G.: Stable isotope disequilibrium between soil bound water and soil bulk water – Implications for estimations of plant water sources, J. Hydrol., 650, 132544, https://doi.org/10.1016/j.jhydrol.2024.132544, 2025.
Yan, Y., Yang, Y., and Dai, Q.: Effects of preferential flow on soil nutrient transport in karst slopes after recultivation, Environ. Res. Lett., 18, 034012, https://doi.org/10.1088/1748-9326/acb8cc, 2023.
Yang, J., Nie, Y., Chen, H., Wang, S., and Wang, K.: Hydraulic properties of karst fractures filled with soils and regolith materials: Implication for their ecohydrological functions, Geoderma, 276, 93–101, https://doi.org/10.1016/j.geoderma.2016.04.024, 2016.
Younger, S. E., Blake, J., Jackson, C. R., and Aubrey, D. P.: δ2H isotopic offsets in xylem water measurements under cryogenic vacuum distillation: Quantifying and correcting wood-water hydrogen exchange influences, Ecohydrology, 17, e2640, https://doi.org/10.1002/eco.2640, 2024.
Zeng, X., Xu, X., Yi, R., Zhong, F., and Zhang, Y.: Sap flow and plant water sources for typical vegetation in a subtropical humid karst area of southwest China, Hydrol. Process., 35, e14090, https://doi.org/10.1002/hyp.14090, 2021.
Zhang, F. and Zhang, C.: Probing water partitioning in unsaturated weathered rock using nuclear magnetic resonance, Geophysics, 86, WB131–WB147, https://doi.org/10.1190/geo2020-0591.1, 2021.
Zhang, J., Wang, J., Chen, J., Song, H., Li, S., Zhao, Y., Tao, J., and Liu, J.: Soil Moisture Determines Horizontal and Vertical Root Extension in the Perennial Grass Lolium perenne L. Growing in Karst Soil, Front. Plant Sci., 10, https://doi.org/10.3389/fpls.2019.00629, 2019a.
Zhang, Y., Zhang, M., Niu, J., Li, H., Xiao, R., Zheng, H., and Bech, J.: Rock fragments and soil hydrological processes: Significance and progress, CATENA, 147, 153–166, https://doi.org/10.1016/j.catena.2016.07.012, 2016.
Zhang, Y., Ge, H., Liu, G., Shen, Y., Chen, H., Wang, J., Liu, D., and Huang, Z.: Experimental Study of Fracturing Fluid Retention in Rough Fractures, Geofluids, 2019, 2603296, https://doi.org/10.1155/2019/2603296, 2019b.
Zhang, Z., Chen, X., Cheng, Q., and Soulsby, C.: Using StorAge Selection (SAS) functions to understand flow paths and age distributions in contrasting karst groundwater systems, J. Hydrol., 602, 126785, https://doi.org/10.1016/j.jhydrol.2021.126785, 2021.
Zhao, Y. and Wang, L.: Insights into the isotopic mismatch between bulk soil water and Salix matsudana Koidz trunk water from root water stable isotope measurements, Hydrol. Earth Syst. Sci., 25, 3975–3989, https://doi.org/10.5194/hess-25-3975-2021, 2021.
Zhao, Z., Deng, J., Zhang, J., and Shen, Y.: Effects of Film-Mulched Rock Outcrops on Rainwater Redistribution and Maize Growth in the Cropland of a Rocky Karst Area, Water, 16, 254, https://doi.org/10.3390/w16020254, 2024.
Zhou, L., Wang, X., Wang, Z., Zhang, X., Chen, C., and Liu, H.: The challenge of soil loss control and vegetation restoration in the karst area of southwestern China, International Soil and Water Conservation Research, 8, 26–34, https://doi.org/10.1016/j.iswcr.2019.12.001, 2020.
Zimmerman, R. W. and Bodvarsson, G. S.: Hydraulic conductivity of rock fractures, Transport Porous Med., 23, 1–30, https://doi.org/10.1007/BF00145263, 1996.
Zwieniecki, M. A. and Newton, M.: Roots growing in rock fissures: Their morphological adaptation, Plant Soil, 172, 181–187, https://doi.org/10.1007/BF00011320, 1995.
Zwieniecki, M. A. and Newton, M.: Water-Holding Characteristics of Metasedimentary Rock in Selected Forest Ecosystems in Southwestern Oregon, Soil Sci. Soc. Am. J., 60, 1578–1582, https://doi.org/10.2136/sssaj1996.03615995006000050042x, 1996.