the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 05 Nov 2025
| 05 Nov 2025
Divergent water balance trajectories under two dominant tree species in montane forest catchment shifting from energy- to water-limitation
Nikol Zelíková
Jitka Toušková
Jiří Kocum
Lukáš Vlček
Miroslav Tesař
Martin Bouda
Vegetation interacts with both soil moisture and atmospheric conditions, contributing to water flow partitioning at the land surface. Therefore, changes in both climate and land cover with vegetation affect the availability of water resources. This study aimed to determine the differential effects of climate change on the soil water regime of two common Central European montane forest types: Norway spruce (Picea abies L.) and European beech (Fagus sylvatica L.). A unique dataset, including 22 years (2000–2021) of measured soil water potentials, was used with a bucket-type soil water balance model to investigate differences in evapotranspiration and groundwater recharge both between the forest types and across years. Results revealed an accelerating transition from a fully energy-limited state towards water-limitation, with evidence of strict water-limitation in recent outlier years, unprecedented in this system. While long-term column-averaged pressure heads indicated drier soil at the spruce site overall, this was driven by the wettest years in the dataset. Seasonal and interannual variability of meteorological conditions drove complex but robust differences between the flow partitioning of the two forest types, which diverged further with increasing water-limitation. Higher snow interception by spruce (27 mm per season) resulted in drier soil below the spruce canopy in the cold season. Higher transpiration by beech (100 mm per season) led to increasingly drier soils over the warm seasons causing lower ground water recharge (34 mm per season). Low summer precipitation inputs exacerbated soil drying under beech more than under to spruce. These suggest that expected trends in regional climate and forest species composition may interact to produce a disproportionate shift of recharge from the summer to the winter season.
- Article
(4293 KB) - Full-text XML
-
Supplement
(909 KB) - BibTeX
- EndNote




Making ecohydrological predictions in a non-stationary state of the Earth system requires detailed process understanding that remains elusive. A major obstacle to advancing process understanding is the lack of long-term observations of variables with direct mechanistic relevance, such as water potential (or hydraulic head). Water potential in soil and plants suffers from a noted information gap despite being key to our understanding of land-atmosphere interactions (Novick et al., 2022). Soil moisture status integrates the fluxes of the entire hydrological cycle and in turn exerts significant control over key Earth system processes (Legates et al., 2011; Humphrey et al., 2021). As water potentials directly drive the soil-plant-atmosphere water flows that are tightly coupled with other land-atmosphere fluxes, addressing this gap offers a promising pathway to resolving major uncertainties in ecosystem fate and functioning (Trugman et al., 2018; Green et al., 2019) during the transition to previously unobserved hydroclimatic regimes. After centuries of relative climatic stability (Brázdil et al., 2022), a clear rise in average and maximum air temperatures has been affecting Central Europe since the last part of the 20th century (Zahradníček et al., 2020). Increased air temperature has induced higher atmospheric water demand contributing to the severity of recent droughts (Možný et al., 2020). Although, long-term annual precipitation sums have not changed in the past (Brázdil et al., 2021) and are not expected to change significantly in near future (Svoboda et al., 2017), the occurrence of seasonal precipitation deficits causing severe soil drought is projected to increase (Hari et al., 2020). Increased water demand combined with seasonally reduced water supply is expected to shift the region from energy- toward water-limitation of evapotranspiration over the coming decades (Denissen et al., 2022).
One of the less well understood consequences of ongoing climatic changes is a shift in forest species composition, which has the potential to further affect water fluxes in the soil-plant-atmosphere system (Maxwell et al., 2018). The two most frequent tree species in central European forests are beech (Fagus sylvatica L.) and spruce (Picea abies L.). As spruce thrives in colder and moisture-rich conditions, its stands are increasingly being replaced by beech (Daněk et al., 2019). This climate-induced transformation of montane forests has potential implications for ecosystem ecohydrological function. Each of these species has distinctive physiological and architectural properties such as leaf morphology and phenology, rooting depth (Jost et al., 2012), xylem structure and function (Tyree and Zimmermann, 2002), or stomatal control during dry periods (Gebhardt et al., 2023). Their specific ecohydrological characteristics and strategies may not only determine their fates under hydroclimatic change but also yield divergent effects on the water balance through contrasting rates of interception (Savenije, 2004), soil water fluxes, water storage dynamics, and thus soil water regimes (Schume et al., 2004).
At present, available studies comparing soil moisture regimes under these common tree species provide ambiguous results due to their limited duration. Schume et al. (2004) and Šípek et al. (2020) reported a stronger drying of the soil profile during the growing season at beech sites. By contrast, Schwärzel et al. (2009), Rötzer et al. (2017), and Kuželková et al. (2024) observed greater soil drying under spruce than under beech. Some of these differences may partly be explained by contrasting soil hydraulic properties at the sites compared. The main limitation shared by such studies; however, is their limited temporal extent. The periods of the observation range from 1 d (e.g., Jost et al., 2012) to several years (Schume et al., 2004; Schwärzel et al., 2009; Zucco et al., 2014; Korres et al., 2015; Huang et al., 2016; Rötzer et al., 2017). The longest periods of analyses so far lasted from 4 to 5 years (Wang et al., 2018; Šípek et al., 2020; Gebhardt et al., 2023). The results of short-term studies are difficult to interpret as they provide only a partial insight into the role of individual water fluxes. They are limited by the variability of climatic conditions during the study period. Moreover, short-term studies cannot capture long-term changes in the characteristics of droughts, such as higher temperatures (Grossiord et al., 2020) and flash droughts (Qing et al., 2022) and therefore their second-order effects via the given species. Hence, the availability of a long-term data series is crucial not only to observe trends, but also as a tool to better understand processes and natural variability in a period of changing climate and land cover (Huntingford et al., 2014; Milly et al., 2015).
This study aims to advance process understanding by disentangling the effects of climate and forest composition on water fluxes as these ecosystems transition from energy- to water-limitation. We focused on the impact of two forest types, monospecific Norway spruce and European beech, on the soil water regime in an experimental montane catchment in Bohemian Forest, Czechia. The study benefits from a unique 22-year-long dataset of measured soil water potential in the two forest types that enables us to make robust interannual comparisons for the first time. Long-term observations of the experimental catchment allow us to impose closure on the hydraulic balance to estimate individual fluxes and to compare the current evapotranspiration regime with previous decades. Together with its depth coverage over the rooting zone in each stand, these advantages allow the present dataset to yield comprehensive insight into the studied forests' ecohydrological function during the ongoing hydroclimatic transition. To reveal how climatic drivers interact with vegetation processes to produce hydrologic flux partitioning, we: (1) analyse seasonal differences in measured soil water potential between the two forest types, (2) estimate the soil water balance components (evapotranspiration and drainage) at the two sites using a process-based soil water balance model, and (3) determine the main climate dependency of the soil water regime under both tree species.
The study is based on extensive field measurements of soil moisture regime and necessary hydrometeorological variables in a Central European montane catchment including spruce and beech covered sites. The water balance of both sites was estimated using the bucket type soil water balance model. The workflow of the study is presented in Supplement (Fig. S1).
2.1 Study site
The Liz experimental catchment, Czechia (49°04′ N, 13°41′ E) (Fig. 1), served as the experimental area for this study. It is located in the Bohemian Forest on the border between Czechia and Germany. The catchment area is approximately 1 km2. Its elevation ranges from 828 m a.s.l. (at the outlet) to 1070 m a.s.l. It is located in the cold region (unit C7 of the Quitt Climatic Classification, Vondráková et al., 2013) of an otherwise humid continental climate (unit Dfb of the Köppen Climatic Classification (Tolasz et al., 2007). During the study period 2000–2021 (and the preceding period of catchment measurements, 1975–1999) it had an average annual air temperature of 7.2 (6.4) °C and an average annual precipitation of approximately 847 (842) mm. The monthly average maximum temperature is 16.5 (15.5) °C in July, and the minimum is −1.9 (−2.3) °C in January. More precipitation arrives during the May–October growing season than the rest of the year: 515.7 (471.2) mm compared with 331.9 (370.9) mm, respectively. Mean annual snow cover duration is 133 (147) d. The annual potential evapotranspiration (PET) determined by the air temperature-based method (Oudin et al., 2005) is 560.7 (521) mm. The annual runoff height from the catchment is approximately 352 (317) mm, representing ∼40 % of the total precipitation.

Figure 1Overview of the experimental site (©CUZK 2024) and soil profiles (© Přemysl Fiala).
Crystalline bedrock in the catchment only allows water circulation in the weathered zone and does not communicate with adjacent catchments, such that the hydrological catchment corresponds fully to the hydrogeological catchment (Hrkal et al., 2009). This observation underpins a fundamental assumption of our modelling framework: that all water from precipitation generates measurable runoff at the gauging station, which is well supported by the hydrogeological survey. The majority of the area is covered by nearly pure spruce forest, with a dominance of 120–140-year-old Norway spruce (Picea abies L.) (>85 % of the canopy cover). In several places, the spruce forest is penetrated by 100–120-year-old beech stands (Fagus sylvatica L.).
Two experimental sites within the Liz experimental catchment were chosen for this study: one with Norway spruce (Picea abies L.) and the other with European beech (Fagus sylvatica L.). The elevation difference between the two sites is approximately 30 m: the spruce site is located at 855–860 m a.s.l., and the beech site is located at 885–890 m a.s.l., both with a slope of 7.5° and an eastern aspect. Both spruce and beech canopies tend to suppress understory vegetation, which was accordingly absent at both sites (Fig. 1). The leaf area index (LAI) was measured throughout the 2022 season on a monthly basis and showed a seasonally stable value with an average of 3.7±0.5 in the spruce site and seasonally variable values in beech ranging from 1.1±0.2 at the beginning and end of growing season (May and September) to 4.7±0.5 in the middle of the growing season. A visual inspection of the root depth distribution (when excavating the soil) revealed that the roots were present only in the upper 40 cm of the spruce site and down to 100 cm of the beech site.
The soil at both sites can be classified as moderately deep loamy sand dystric Cambisol (IUSS Working Group WRB, 2015), with an average soil depth of approximately 100 cm. The percentages of sand-silt-clay fractions are 73.2 %–24.2 %–2.6 % at the spruce-covered site and 80.2 %–18.1 %–1.7 % at the beech-covered site. The soil water permeability is relatively high ranging from 518 cm d−1 at the bottom of the soil profile to 1700 cm d−1 in the topsoil horizon. The humus A horizon (0–10 cm), together with surface organic horizon O (5–10 cm thick at beech stand and 10–15 cm at spruce stand), is followed by a Bvs/v horizon (down to 50 cm at beech site and to 30 cm at spruce site) and finally by a BvC horizon with a significant amount of larger than sandy particles (>50 %). Both soil profiles are presented in Fig. 1.
2.2 Field measurements
The meteorological variables used in this study were air temperature (Fiedler RV12/RK5, Czech Republic) and precipitation (Meteoservis MRW 500, Czech Republic), which were measured at 10 min intervals during the entire twenty-two-year period (2000–2021). The meteorological station is located circa 400 m away from two experimental plots outside the forest. Moreover, the experimental catchment is instrumented with discharge and groundwater level measurements. Discharge was also measured at the 10 min time step, and the groundwater level was recorded manually every week throughout the entire investigated period. Average daily air temperatures, precipitation sums and discharges were collected from 1975. The snow water equivalent (SWE) was measured manually three times per week since 2000. Soil water potential data were acquired from permanently installed soil tensiometers (Adolf Thies GmbH, Germany, see Fig. S2) measuring pressure heads at five depths (15, 30, 45, 60 and 90 cm). Soil water potentials were recorded manually three times a week during the growing season (mid-May to mid-October) from 2000 to 2021. The measuring range of these tensiometers included pressure heads ranging from 0 to −865 cm (−85 kPa). Up to four tensiometers were available for each measuring depth at each site over the entire measurement period (at least 2 measurements 93 % of the time), and we used their average for a particular depth as the site-representative value. Given the fully closed, even, monospecific canopies at our sites, the representativeness of the measurements was ensured by avoiding placing sensors at micro-sites subject to preferential flows. We used measurement points representing average site slope and distance between the trees ( m from tree in spruce/beech forest when the average distance in between two adjacent trees is m). This resulted in same order of spatial variability of LAI (coefficient of variation was % for spruce and beech) and soil moisture (coefficient of variation was % for spruce and beech) in both forests and good correspondence of soil water potentials with another three profiles equipped with UMS T8 tensiometers located nearby (Šípek et al., 2020). The average soil column pressure head was estimated as a weighted mean of five soil layers (each represented by one measurement depth). The soil profile was considered to have a uniform depth of 100 cm. The measured pressure heads were used to determine differences in soil water regimes between the stands, as they better demonstrate the differences in soil water energy states during dry conditions, which were of interest to the study.
2.3 Soil water balance model
The conceptual model used in this study was a modified form of the soil water balance model (SWBM), developed by Brocca et al. (2008, 2014). The bucket-type of the model was used as (1) it is sufficient to answer questions posed (soil column water balance) without adding more complexity, (2) it uses “Feddes” type of equation for the estimation of plant water use, (3) it is more convenient for the simulation of longer periods, (4) the soil column is represented by one unified domain with column average soil hydraulic properties, which is beneficial especially when the soil encompasses a lot of rock fragments. Moreover, several widely used hydrological models use similar bucket/reservoir modelling approaches for the determination of soil water regimes (e.g., the Soil Water Assessment Tool (Arnold et al., 2012), the HBV model (Seibert and Vis, 2012) or the VIC model (Liang et al., 1994)). The modification for this study is based on the replacement of the infiltration parameter (the Green-Ampt equation) by throughfall (PTF), as surface runoff is not generated in the experimental catchment and all water directly infiltrates into the soil. Therefore, the following soil water balance Eq. (1) was used:
where θ(t) is the average volumetric soil water content at a day (t), PTF(t) is the throughfall (mm d−1), S(t) is the actual evapotranspiration rate (mm d−1) and D(t) is the drainage rate (mm d−1). The Eq. (2) for PTF(t) is given as:
where POAR represents the measured open area precipitation (mm d−1) and PINT is the estimated interception (mm d−1) for a given location. Spruce interception in the summer season (May to October) was estimated based on the deduction of the interception capacity from every single precipitation event. The interception capacity of 2.2 mm was derived by Kofroňová et al. (2021) for the same experimental site. In the case of beech stands, the summer interception capacity was calculated using a general formula by von-Hoyningen-Huene (1983) and Braden (1985) applying seasonal variation in the leaf area index (LAI):
where a is an empirical coefficient (–) and b is the soil cover fraction ( (–). Daily values of LAI were acquired from linear interpolation between monthly measured values (May–September) conducted by a LI-COR 2000 Plant Analyser in 2022 (Toušková et al., unpublished results). The calibration of a parameter was performed so that the fraction of intercepted precipitation was allowed to range between 15 and 20 %, which is an ordinary interception loss of beech canopies (Gerrits et al., 2010). For the winter season (November to April), linear regression functions linking open area snow water equivalent to that below the forest canopy were used (Šípek and Tesař, 2014). The regression equations are based on the measured snow water equivalents in the forest openings and below the spruce (Eq. 4) and beech (Eq. 5) canopies for a period of ten years and are in the form:
where SWEOAR is the snow water equivalent (mm d−1) in the open area and SWETF is the snow water equivalent under the forest canopy (mm d−1).
Potential evapotranspiration (PET) was estimated using the Oudin et al. (2005) approach, which offers reliable estimates of PET for long-term water balance studies in the Central European region (Toušková et al., 2025). This approach provided a consistent PET estimate based on data available for the entire observation period (1975–2021), whereas data needed for more sophisticated approaches are not available for the first decades. The actual evapotranspiration (AET) was found as the sum of PINT and soil evapotranspiration rate S (comprising soil evaporation and plant transpiration) was then estimated based on the linear decrease in its potential rate with decreasing effective soil water content as proposed by Feddes and Rijtema (1972) according to the following Eqs. (6) and (7):
where PET is the potential evapotranspiration (mm d−1), Θe is the effective water content (–), and Θr,s are the residual and saturated soil water contents (mm), respectively and θ(t−1) is modelled volumetric water content at previous day (t−1). The drainage component D(t)is a nonlinear function of Θe:
where Ks is the saturated hydraulic conductivity (mm d−1) and λ is the pore size distribution index (–) linked to the textural structure of the soil layer, which was set to 0.5. In this case, the flow is assumed to be gravity driven, with drainage consisting of deep percolation.
The original SWBM does not include a snow module; hence, snow accumulation and snowmelt had to be considered first, as the experimental catchment lies in an area with regular snow cover. The degree-day method (Gupta, 2001) was chosen for this purpose because it has been proven to be efficient in the Central Europe (Girons Lopez et al., 2020).
2.4 Model parameterisation, validation, and forward simulation
The model was calibrated with the genetic algorithm in two separate steps: one focused on the additional snow module and the second on the original SWBM parameters using fixed values of snow parameters from the first step. In each case, the RMSE of the model response variable (snow water equivalent and soil water content, respectively) was used as the objective function. All model parameters which were subject of calibration are described in Table 1. Four parameters of the snow module were calibrated separately for each winter season so that the input for the soil water model was as accurate as possible. The remaining model parameters were calibrated against the soil water content at both the beech and spruce sites. To obtain soil water content for calibration, the measured pressure heads were used to calculate the volumetric soil water content by means of the Van Genuchten (1980) function. The function parameters were retrieved from the measured retention curves specific for each site and depth (see Table S1 in the Supplement). For more information about the determination of the soil water retention curves, we refer to Šípek et al. (2020). In addition to the minimisation of the RMSE, the model was calibrated with three boundary conditions: (1) simulated drainage from both sites must be approximately 360 mm yr−1, which is a value obtained from the long-term measured runoff from the area, (2) higher reported transpiration of beech (Brinkmann et al., 2016; Gebhardt et al., 2023) and (3) beech summer interception loss will be within 15 %–20 % of the open area rainfall, which corresponds to the range reported by Gerrits et al. (2010).
To evaluate model fit, we first split the period of interest into 4 sub-periods for cross-validation, each covering 5 years (2000–2004, 2005–2009, 2010–2014, 2015–2019) and calibrated the model separately for each of these periods using the remaining three sub-periods as unseen datasets for cross-validation. In each case, we constrained drainage to fit the measured runoff by omitting the parameter sets resulting in differences in runoff volume higher than 5 %. Model error in cross-validation was on the same order as measurement error (max. RMSE <3 %, see Fig. S3) and parameters did not change substantially when all sub-periods were used. We thus chose to calibrate a single set of model parameters for the entire period so that the water balance (i.e., discharge) could be maintained as close as possible to the measured long-term mean. Using mean drainage estimated from the water balance for the whole period as a constraint led to only minor deterioration of the objective function compared with parameters fitted to the separate four 5-year sub-periods.
Following validation, we thus only used the model fitted to the entire period of available data (2000–2021). Besides the model run in the period of available soil water potential measurements (2000–2021), the calibrated model was run also from 1975 to 1999 when soil water potential measurements were not available in order to quantify annual AET and runoff for the period spanning to the beginning of the meteorological measurements.
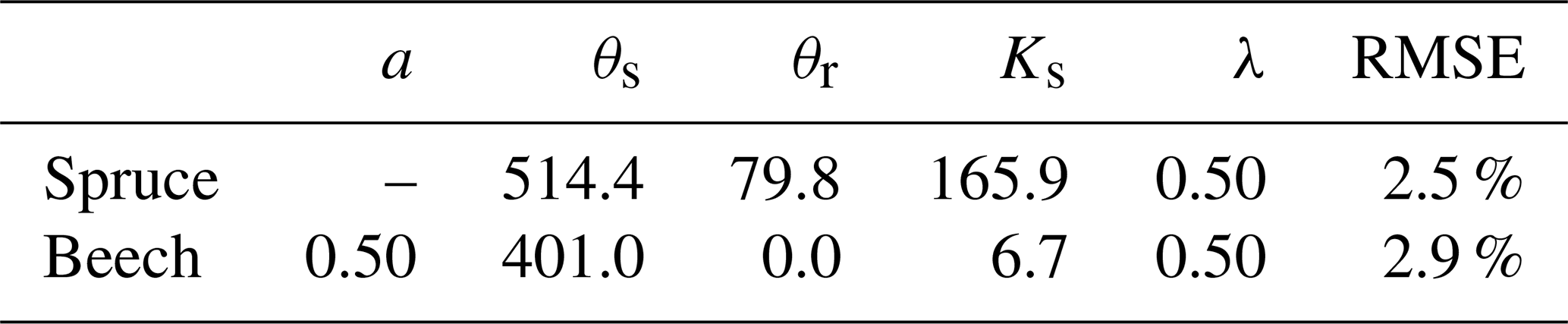
Table 1Model parameters. SWE stands for measured snow water equivalent and SWC for soil water content.
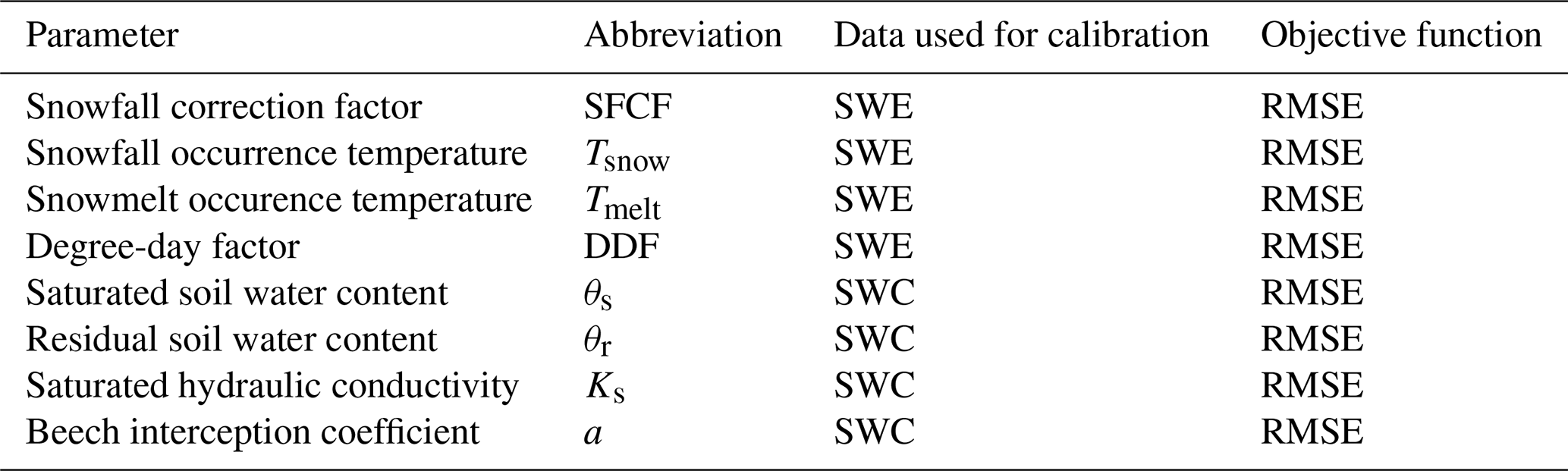
2.5 Analysis
Meteorological data and soil water contents trend analysis was conducted using trend-free pre-whitening Mann-Kendall approach (Yue et al., 2002). We also calculated annual and 5-year evaporative ratio () and aridity index () values from our model results to evaluate energy versus water limitation of our sites using the Budyko framework (Renner et al., 2014; Mianabadi et al., 2020). For the analysis of the vertical distributions of pressure heads we utilized a principle of flow-duration curves describing the fraction of time that the magnitude of a given variable is exceeded (Dingman, 2015).
3.1 Analysis of the measured soil water regime
The climate conditions of all investigated summer and winter seasons are depicted in Fig. 2. One wet and one dry year were chosen to demonstrate differences among pressure heads between the spruce and beech sites influenced by extreme meteorological conditions (Fig. 3). Specific years were then categorized according to soil wetness régime based on the seasonal distribution of measured pressure heads from May to August.
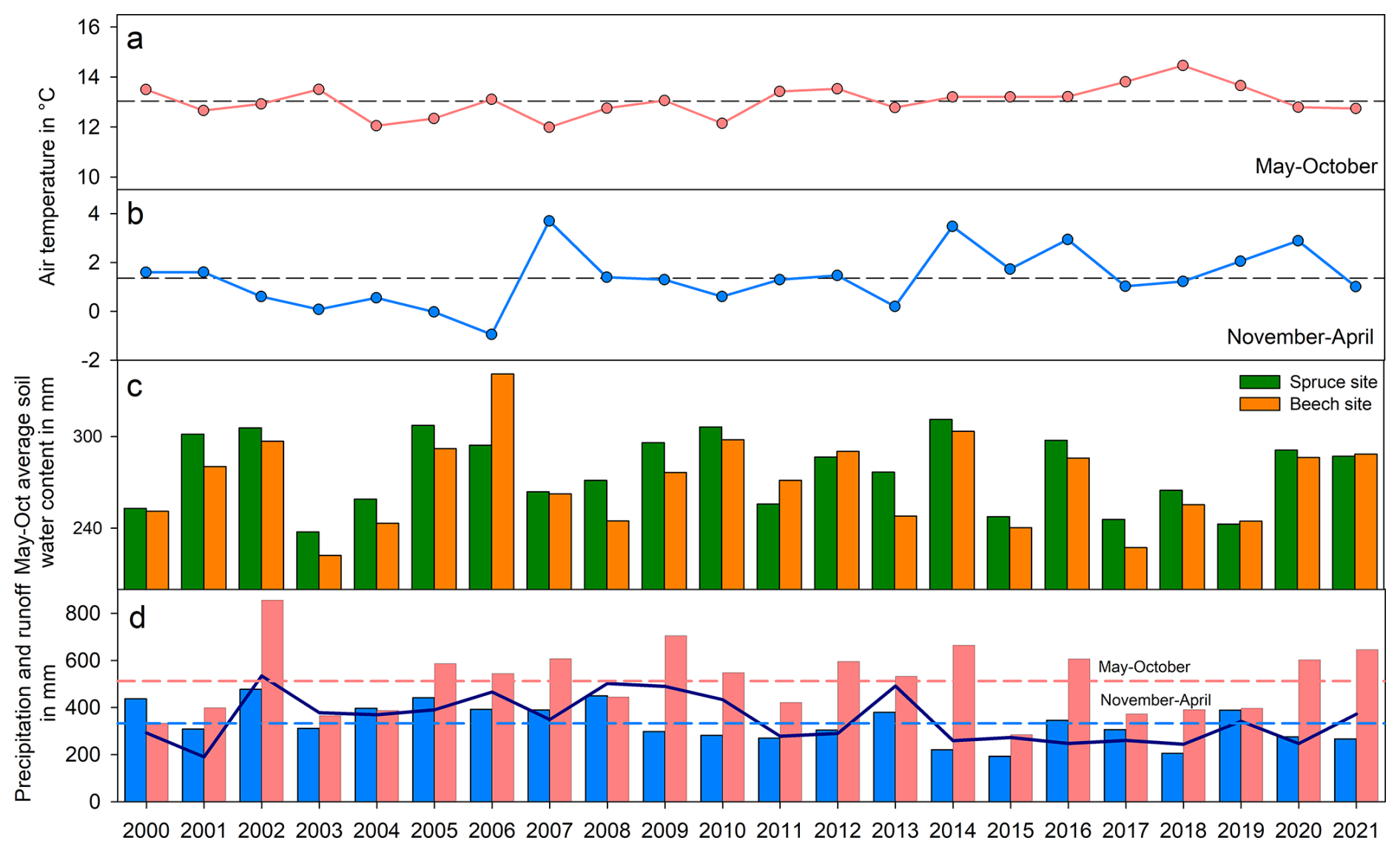
Figure 2Average air temperatures in summer (a) and winter seasons (b), May–October average soil water content (c) and precipitation sums and runoff (d). The red columns represent the summer seasons (May–October), and the blue columns represent the winter seasons (November–April), dashed lines represent season average values, and dark blue line represents runoff.
3.1.1 Vertical distribution of pressure heads
Pressure-head values were higher at the beech site, with a long-term median of −155 cm compared to −255 cm for the spruce site. However, despite the higher median pressure-head values recorded at the beech site, the occurrence of low pressure-heads was more frequent here as reflected by higher exceedance of pressure values lower than −400 cm from the depth of 30 cm and deeper (Fig. 3). Differences in the vertical distribution of pressure heads were visible, namely, in the topsoil layer (depth of 0–15 cm), where soil under spruce reached permanently lower pressure head values than that under beech. The overall depth distribution of the pressure heads was more uniform under spruce – documented with flatter slope of curve describing the exceedance of pressure heads in all depths (Fig. 3). In contrast, the pressure head depth distribution under beech trees exhibited greater propensity to drying, especially in the bottom soil layers. The slope of exceedance curve is steeper namely between pressure heads of −200 and −400 cm. As the soil gets drier then the soil water potential is lower under beech. The beech site, despite having higher pressure heads on average, was therefore more susceptible to more intensive drying than the spruce site.
The differences between the beech and spruce site were less pronounced during the wet years (e.g., year 2020 represented by long-dashed lines in Fig. 3) but the soil under beech was noticeably drier in dry years (see example dry year 2015 represented by short-dashed lines in Fig. 3). Although the differences among the sites were small in wet years, lower pressure heads were observed at the spruce site at all depths. In contrast, during the dry year of 2015, the soil under spruce site was wetter (reached a higher column average median pressure head) than at the beech site even in the top soil layer (depth of down to 15 cm). Below the depth of 45 cm the pressure head of −850 cm was exceeded in more than 50 % of records under beach and only up to 10 % in the case of soil under spruce canopy. Hence, the differences in pressure heads might be even greater, as the tensiometer data reached their limit more frequently at the beech site than at the spruce site; thus, even lower pressure heads were likely to occur at the beech site. If the number of dry years increase in the future, the soil under beech will therefore become drier during the growing seasons.
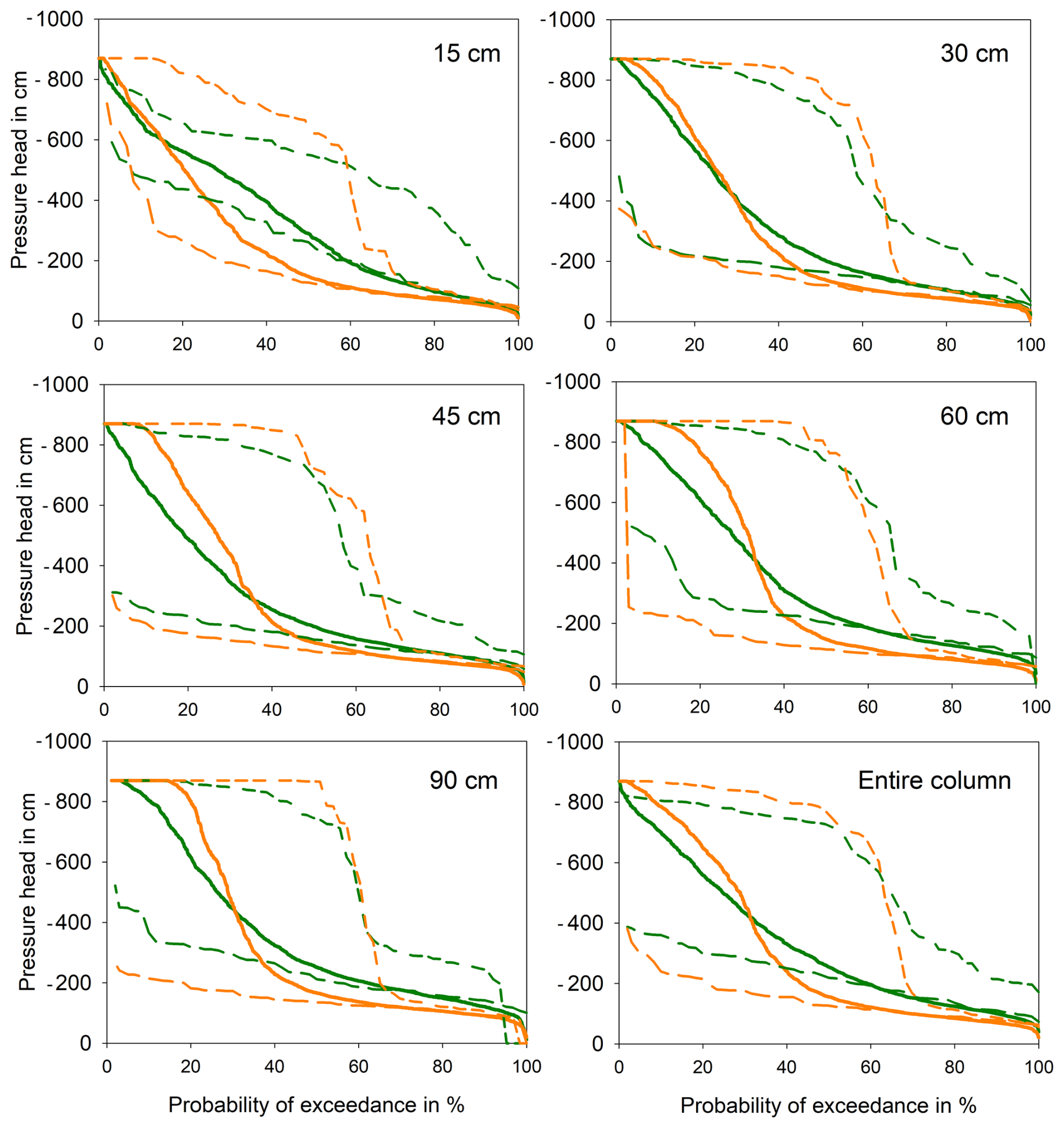
Figure 3Exceedance probabilities of pressure head for particular depths for averaged the entire period (thick solid lines), dry season 2015 (short dashed lines) and wet season 2020 (long dashed lines). Green colour represents spruce and orange beech forest.
3.1.2 Soil wetness trend and categories
We found significant negative trends in both daily soil moisture time-series, 0.7 mm yr−1 in beech (p value 0.001), 0.2 mm yr−1 in spruce (p value 0.0015), documenting gradual changes in soil water regime which correspond to the increasing occurrence of water limited seasons. Looking closer, we divided years into four soil wetness categories based on the typical seasonal development of their measured pressure heads.
We have used four categories of soil moisture regime for further analysis:
-
category A – spruce retained lower pressure heads throughout most of the season
-
category B – only one single event when the beech site attained lower pressure heads than spruce
-
category C – the pressure head decreased more pronouncedly at the beech site for a significant part of the summer season
-
category D – refers to the seasons when the tensiometer measurement limit of −865 cm was reached (mostly at the beech site)
The evolution of average pressure heads for each month of the summer season over the measured period (2000–2021) is depicted in Fig. 4. At both sites, a similar pattern of decreasing pressure heads from the onset of the summer season can be observed. However, there are noticeable differences between the two sites. At the beginning of every summer season (May), the spruce site reached lower pressure head values than did the beech site (the average difference in pressure heads was 130 cm). Typically, as the season progresses, the pressure heads at the beech site decrease more than those at the spruce site. However, this was not valid for the wet seasons of 2002, 2005, 2020, and 2021, when spruce retained lower pressure heads throughout most of the season (see Fig. 4), as no precipitation deficit was observed (category A). For those seasons, the difference between the two sites was negligible, with their average values fluctuating between −100 and −200 cm. In the other few years, when above average precipitation seasonal sums were reached (category B, including the years 2006, 2010, 2012, 2014, and 2016), there was only one single event when the beech site reached lower pressure heads (below −400 cm), which was usually ended by rainfall higher than 50 mm d−1. In contrast, in the periods with below average precipitation, the pressure head decreased more pronouncedly at the beech site for a significant part of the summer season (category C included, e.g., years 2007, 2012 or 2019, as shown in Fig. 4). With even more prominent precipitation deficits (in 2003, 2008, 2015 and 2017), the beech site was the first and often only site to reach the tensiometer measurement limit of −865 cm (category D) – up to ten times more often than the spruce site, especially in the bottom soil layers. Real pressure head values were likely significantly lower. As lower pressure heads cannot be recorded at the beech site with tensiometer measurements (measuring limit was reached) and pressure heads at the spruce site only seldom approached this limit, the differences between both sites were higher than documented by sensors. The effect on our analysis was likely insignificant as the implied differences in the amounts of water retained would be rather small. By the end of the season, pressure head values slowly increased, with beech still maintaining lower pressure head values than spruce.
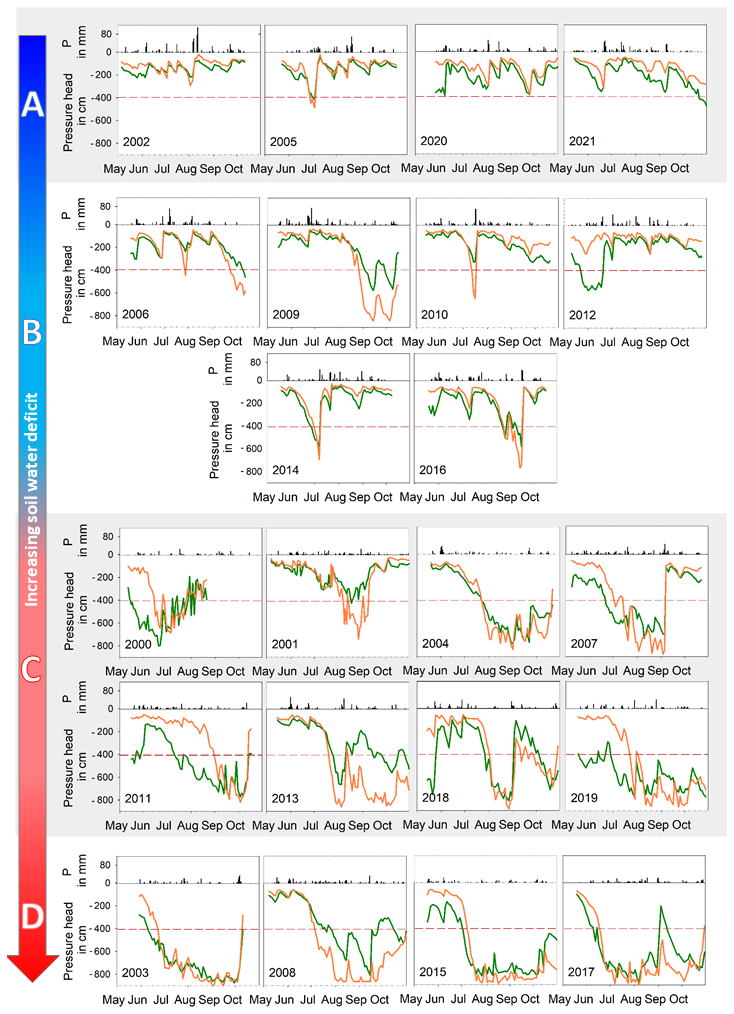
Figure 4Daily precipitation (P) (black columns) and soil column average pressure heads at beech (orange line) and spruce (green line) sites in all investigated years divided into four wetness categories (A–D) defined by pressure head values. The red dashed line represents the pressure head of −400 cm used for the division of categories A and B.
3.2 Modelling of evapotranspiration and drainage
3.2.1 Model calibration result
The modified SWBM model was used to obtain evapotranspiration and drainage fluxes over a period of twenty-two years (2000–2021) at both spruce and beech sites. The mean RMSE values (2000–2021) for the snow module were 7.1 mm (beech) and 9.5 mm (spruce), which are in accordance with Šípek and Tesař (2017), who modelled snow cover dynamics from 2009 to 2014 and reached an RMSE value of 9.1 mm in a spruce stand. An example of the modelled cumulative snow precipitation fitted to the measured SWE is shown in Fig. S4a.
The resulting mean RMSE (2000–2021) were 2.5 % and 2.9 % for the spruce and beech sites, respectively. The modelled long-term drainage was 369 mm yr−1 for beech and 365 mm yr−1 for spruce. The average annual discharge for the experimental Liz catchment was 360 mm, which was very close to the modelled values. The final parameters of the SWBM (θs,θr, Ks,λ) for each site are documented in Table 2. Examples of modelled and observed volumetric water contents are depicted in the Supplement (Fig. S4b).
3.2.2 Simulated Water balance
The total actual evapotranspiration (AET; encompassing transpiration and soil evaporation (S) and interception (Pint)) and drainage attain similar values at both plots on average. The total average AET is approximately 540 mm yr−1, and the drainage is between 350 and 360 mm yr−1 (Table 3). The beech reaches on average almost 80 mm yr−1 more S than the spruce stand, on the other hand, the evaporation from the interception storage in the spruce stand exceeds that of the beech stand to the same extent. The resulting AET values therefore do not differ greatly from each other because S and interception tend to compensate for each other between stands, which is hence also reflected in similar drainage.
Even though the winter seasons are characterised by lower precipitation sums than the summer seasons (approximately of the annual precipitation), the spruce forest had, on average, a higher rate of interception (133 mm per season) due to defoliated beech forest (106 mm per season) (Table 3). From the AET perspective, the difference in interception is further raised by slightly higher transpiration and soil evaporation under the spruce canopy (11 mm per season). Nevertheless, the interception rate and winter transpiration at the spruce site resulted in a lower amount of water available for infiltration and therefore a lower modelled soil water content during the winter months. The drier soil in spruce forests regularly represents an initial condition for the summer season. A higher soil water content below the beech canopy was a reason for higher modelled drainage during the winter season at the beech site (by 34 mm per season on average).
Table 3Modelled soil water balance components (mm) at the spruce and beech sites. S represents transpiration and soil evaporation from the soil column. AET stands for actual evapotranspiration.
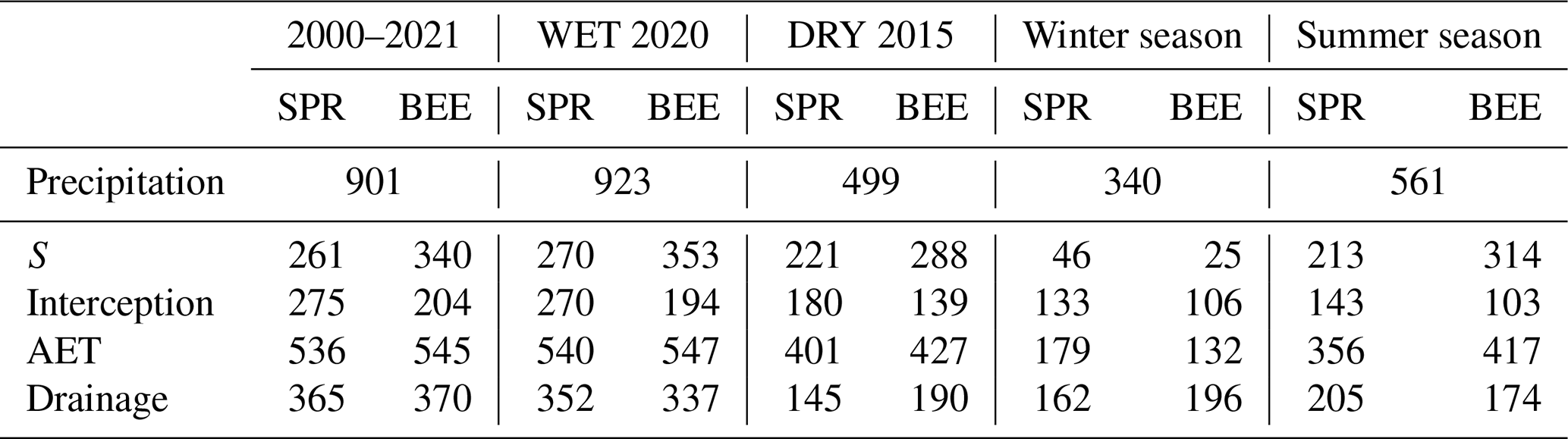
In the summer, transpiration flux significantly affected the water balance at both sites but it was noticeably higher in the beech forest (see Table 3). The interception pattern of both stands was preserved, with spruce having higher interception (143 mm per season) than beech (103 mm per season). The differences in the soil water content were therefore caused by the transpiration in the beech stands (by 101 mm per season on average). Hence, soil under spruce trees retained (with the ongoing summer season) more water than soil under beech trees, where soil moisture was more effectively used for higher transpiration of beech trees, especially during dry spells. The wetter soil under spruce (in the majority of summer seasons) resulted in higher drainage by 34 mm per season on average.
3.3 Interannual comparison of climatic drivers of seasonal soil water regime and soil water fluxes
Figure 5 shows the relative rankings of individual study years according to snow cover duration, air temperature, summer precipitation (May–October), and their classification into the four wetness categories according to the resulting pressure head dynamics, shown in Fig. 4.
The dominant factor controlling the soil water regime in the growing season was the amount of summer precipitation. A significant soil moisture deficit could develop even following a winter with abundant snow. Figure 5 clearly shows the direct link between pressure head and summer precipitation, where lower pressure heads are linked mainly to years with lower seasonal precipitation, and higher pressure heads are linked to years with abundant precipitation. The correlation coefficient between summer precipitation and soil moisture regime category was 0.80 (significant at 0.05 probability level). Two marginal categories (A and D) were always linked to specific climatic conditions (see Fig. 5). Category A, denoting wet soil (hence small differences between beech and spruce sites), was always determined by above average precipitation amounts and below average air temperatures observed in the summer season. Category D, representing the very dry soil moisture regime, was always accompanied by low observed precipitation amounts in the summer season. Two middle categories (wetter B and drier C) tend to be connected primarily with above (in the case of B) and below (category C) average precipitation sums. The influence of preceding winter snow cover and summer season air temperatures was ambiguous, as seen in the frequently strongly mismatched placement of particular seasons along these axes in Fig. 5, compared to the resulting soil wetness category. The correlation coefficient with soil moisture regime were 0.30 and 0.08 for summer air temperature and snow cover duration, respectively. Higher correlation coefficient was also observed for the summer vapour pressure deficit (VPD) attaining the value of 0.61 (not shown in the Fig. 5).
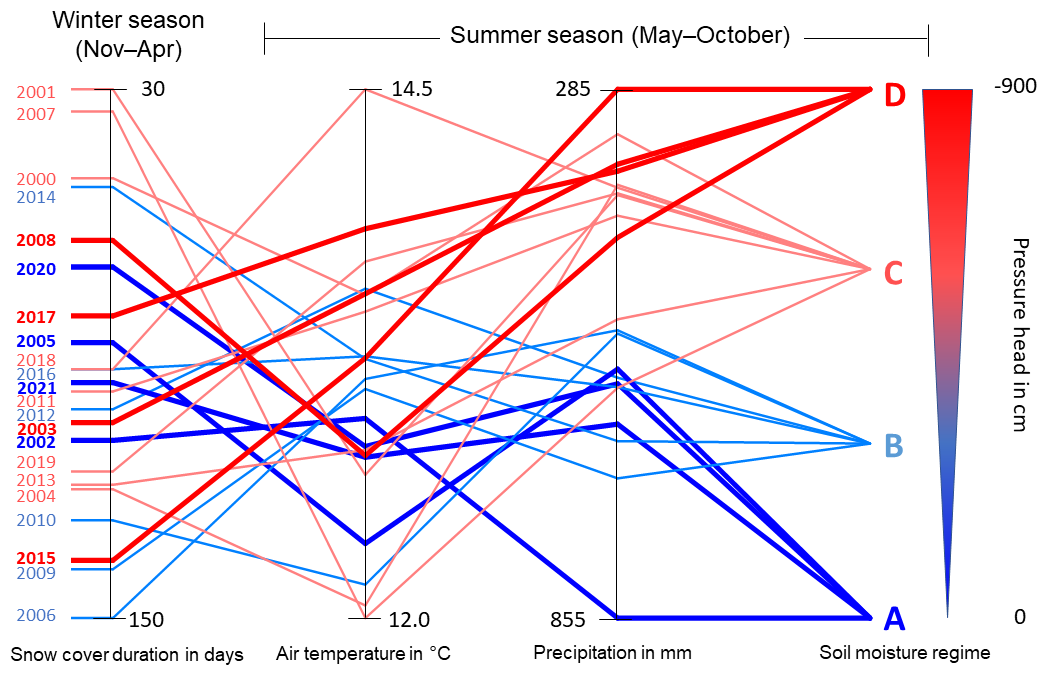
Figure 5Average air temperature and precipitation sums for each summer season (represented by one horizontal line) encompassing the preceding winter snow cover duration. Each season is ultimately linked to a specific wetness category (A–D), as shown in Fig. 4.
The most pronounced deviations from the observed link between summer precipitation sums and the soil moisture regime were in the 2013 and 2007 seasons, with above average precipitation but a drier soil moisture regime. This was caused by a near absence of snow cover observed in the winter of 2006/2007, accompanied by the highest recorded winter air temperature (Fig. 2), and by the extreme floods in 2013, when the catchment received of all summer precipitation in June but saw below average precipitation amounts during the rest of the season. These two factors caused a drier soil moisture regime even when above average precipitation sums were recorded. These results therefore document how different rainfall conditions influence the development of soil moisture content and the different behaviours of beech and spruce in growing season (Fig. 4).
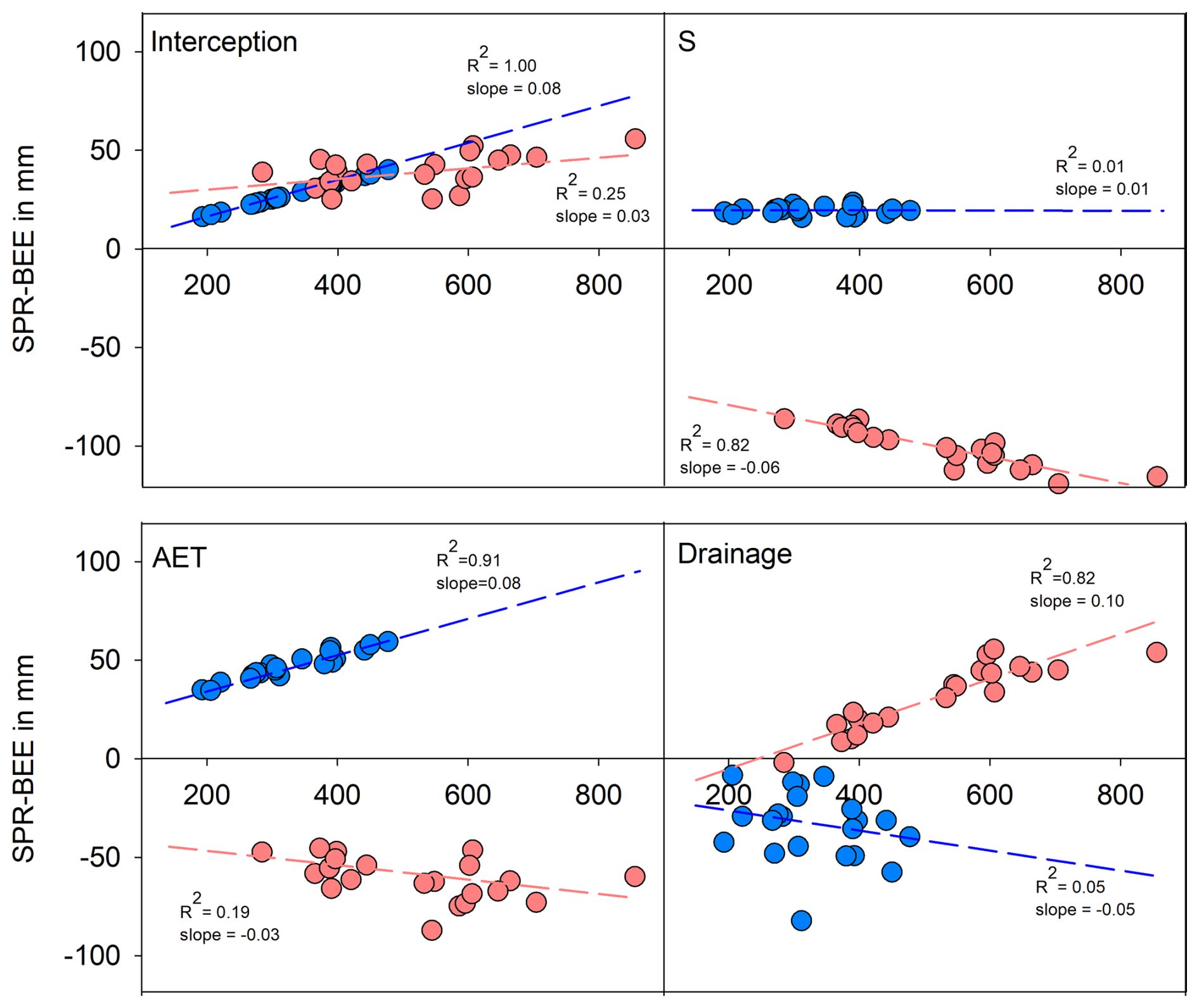
Figure 6Differences between spruce and beech modelled soil water fluxes (AET, D) during summer (May to October, orange colour) and winter (November to April, blue colour) in relation to precipitation. AET can also be divided into INT and S (upper panel).
Seasonal precipitation also had a major influence on the differences between beech and spruce sites in particular water fluxes (Fig. 6). Differences in all fluxes could be positively or negatively related to seasonal precipitation sums with the exception of winter transpiration and soil evaporation (S). The differences in winter and summer interception, winter actual evapotranspiration (governed mainly by interception) and summer drainage increased with increasing precipitation. By contrast, summer transpiration and soil evaporation, summer actual evapotranspiration (governed by transpiration) and winter drainage were negatively related to precipitation sums. The largest absolute differences in water fluxes between the stands were recorded during wet summer seasons. The most pronounced discrepancies were in the rates of transpiration and soil evaporation (higher for beech plots; up to 120 mm per season), summer interception (higher for spruce plots; up to 55 mm per season) and drainage (higher for spruce plots; up to 55 mm per season). The lowest differences occurred during the dry winter seasons. The differences in the winter seasons were generally less prominent, usually below 40 mm per season.
4.1 Transition from energy and water limitation
The studied catchment falls within a montane system classically thought of as energy-limited not just under “baseline” (1961–1990) climate but over previous millennia (Schafstall et al., 2024). Our results show gradual soil drying following an accelerating shift in the balance between atmospheric water supply and demand (Fig. 7). The transition from energy- toward water-limitation predicted for the coming decades (Denissen et al., 2022) is in fact already apparent over our measurement period. Incipient water limitation at the annual-scale was first observed for the drought year 2003 and four times since, with entirely unprecedented examples of outright water-limitation over the years 2015 and 2018. Our dataset thus offers some of the first observations of the hydrologic functioning of these previously cold and humid montane forest types under water limitation.
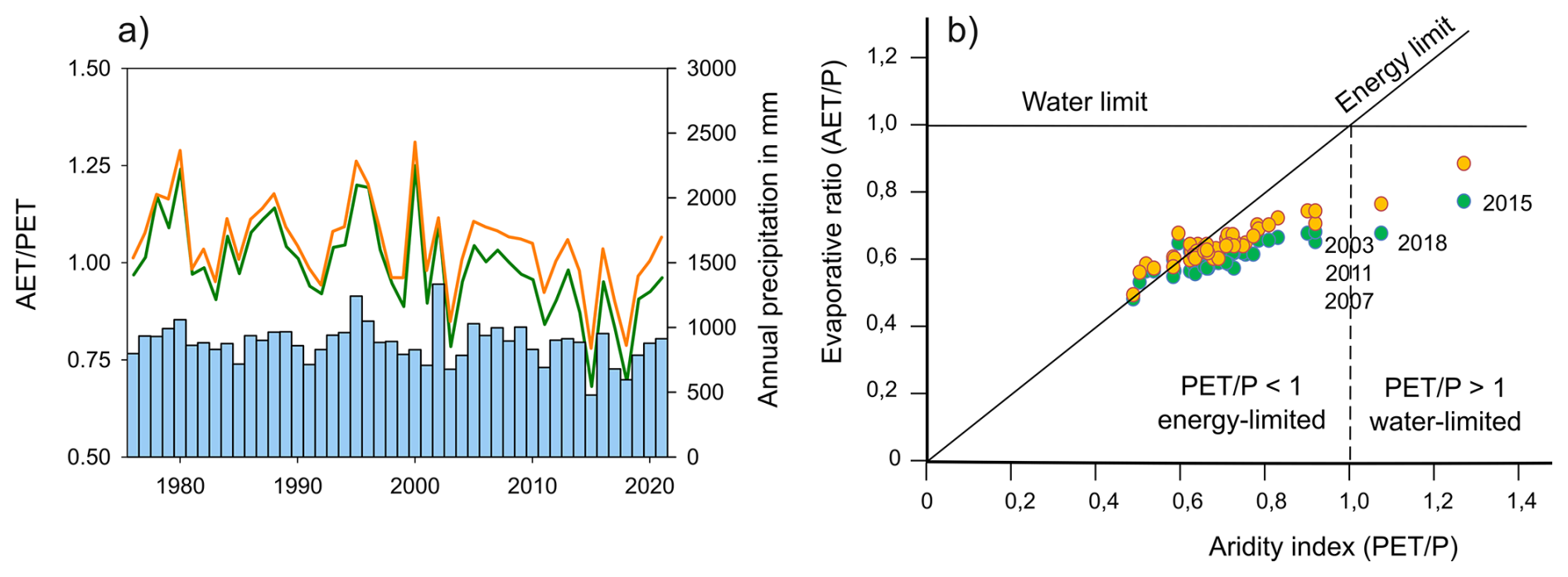
Figure 7Long-term evolution of ration of AET to PET ratio (1975–2021) in the comparison with annual precipitation sums (a), and ratios of actual and potential evapotranspiration to precipitation from the experimental watershed covering the period 2000 to 2021 shown within the Budyko curve reference frame (b). Green lines/points represent the spruce site and orange lines/points beech. Ratios of AET/PET higher than 1.0 are given by the winter snow interception as its estimation is not based on the PET.
We found that increased water limitation enhances differences in annual evaporative ratio between beech and spruce forest, indicating divergence of their water balance in a drier climate. The differences in soil moisture were strongly dependent on the seasonal precipitation sums. Even though, observed trends in the catchment over the period 1975–2021 show that significantly increasing annual atmospheric demand (PET) (slope 1.6 mm yr−1, p value ) rather than insignificant changes in precipitation supply (P) drives increased aridity over the long term, the differences in flux partitioning in the driest years were strongly dependent on the seasonal precipitation sums (Figs. 5–6). With increasing water limitation, the trend of atmospheric demand will cease to exert direct control over the water balance (P−AET), while interactions between seasonal precipitation patterns and vegetation processes will become increasingly important drivers.
4.2 Vegetation and climate interactions in the soil moisture regime
Our unique 22-year long dataset of measured soil water potentials, air temperatures and precipitation sums enabled robust comparisons of the soil water regime between dry and wet years (Fig. 8), allowing modelled soil water fluxes under beech and spruce canopy to reveal the interactions between forest cover, climate, and soil moisture. Differences in winter soil moisture regime were determined mainly by the higher interception of the spruce canopy, which resulted in higher pressure heads under beech causing more drainage compared to the spruce site. When the precipitation in the following summer season was high, only minor differences in pressure heads were recorded between stands, even though the spruce site maintained slightly lower pressure heads throughout most of the season (as a winter season legacy effect). The resulting differences remained small as the higher interception of spruce did not exceed the higher rate of transpiration of beech in the growing season.
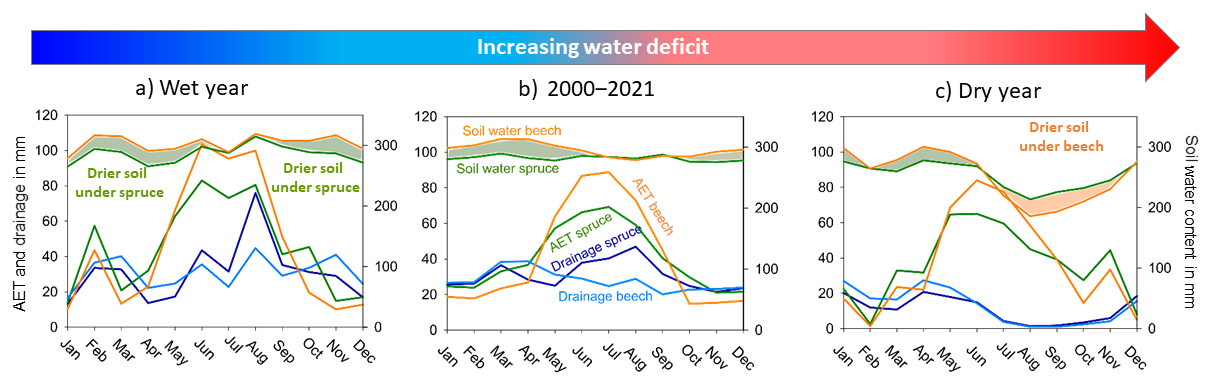
Figure 8A monthly water budget under spruce and beech canopy in wet (2020) and dry (2015) year and its overall averages.
As the growing season advanced, transpiration became an increasingly important factor in the soil moisture regime. The balance between interception and transpiration and soil evaporation resulted in greater drainage under the spruce canopy. In seasons with prominent precipitation deficits (Fig. 8c), the soil at the beech site consistently dried out more than at the spruce site. This can be explained by species-specific plant hydraulic traits. Beech has a wider and deeper rooting pattern and thus soil volume and water in its root zone, especially at greater depths (Čermák et al., 1995; Schwärzel et al., 2009; Gebauer et al., 2012). Beech also has a greater tissue-specific hydraulic conductance due to favourable anatomical and morphological traits, allowing it to supply leaves with water more efficiently at a given root-zone water potential (Tyree and Zimmermann, 2002). As a result, beech behaves more anisohydrically, maintaining transpiration rates in the face of drier soils, in contrast to the more isohydric spruce, whose lower ability to supply water to its foliage requires it to restrict transpiration earlier as the soil dries out (Čermák et al., 1995; Zweifel et al., 2002; Schume et al., 2004; Hochberg et al., 2018; Gebhardt et al., 2023). Schwärzel et al. (2009) and Floriancic et al. (2022) reported higher evaporation from soil and litter under beech stands compared to spruce. Additional factors possibly affecting differences in soil water regimes include lateral flow, which is reportedly more common at beech sites (Jost et al., 2012), and root water redistribution (Burgess et al., 1998). In dry summers, the drainage remained higher under spruce canopy, although the difference between the stands decreased as the difference between interception and transpiration declined.
Robust interannual comparisons of the soil moisture regime under beech and spruce canopies integrated over the entire soil column allow this study to resolve the contradictory results of previous work limited in scope of over space or time. While Schume et al. (2004) and Šípek et al. (2020) observed drier soil during the growing season under a beech canopy, Schwärzel et al. (2009) found the opposite. In the latter case the more prominent drying under spruce was attributed to the nonuniform and rocky soil compared to beech site. Rötzer et al. (2017) and Kuželková et al. (2024) also reported drier soil under spruce but these studies covered only the upper part of the soil profile (0–30 cm). Viewed over two decades and the entire soil profile, the contrasting soil moisture regimes of individual studies prove to be precipitation-driven while differences between the forest types are dominated by depths of 30 cm and more, where the greatest differences arise. The latter finding highlights the need for soil moisture measurement at greater depths, which are too often neglected.
We also found a surprising trend of intra-annual precipitation redistribution in the catchment since 1975. Our observations show significantly decreasing winter (slope −1.7 mm yr−1, p value 0.061) and insignificantly increasing summer P (slope 1.7 mm yr−1, p value 0.24), which is entirely contrary to prevailing expectations based on climate model predictions (Kyselý et al., 2011). Given that seasonal P sums interact with vegetation processes to affect the overall water balance, the actual direction of this trend will not only determine when water arrives in the system but also how it is partitioned.
A precipitation shift in either direction would reinforce the ecohydrological differences between the two forest types. With a shift to winter precipitation, differences in summer transpiration would be abated by lower growing-season input and groundwater recharge would become increasingly reliant on deciduous forest due to its low winter interception. By contrast, a shift to summer precipitation would decrease ET from winter interception by evergreen forest and increase the importance of montane spruce forest to recharge. The rates of groundwater recharge under the two forest types will thus continue to diverge under increased water limitation in either precipitation seasonality scenario.
Further developments in the forest species composition of montane catchments is also likely to play a role. Given the present dominance of spruce, the precipitation seasonality trend in our catchment is consistent with groundwater recharge shifting to the summer and offsetting overall drying somewhat. Increasing representation of beech would exacerbate higher atmospheric demand, given their ability to consume soil water even during drought periods. A combined trend of wetter winters and increasing representation of beech trees in Central Europe, would lead to even higher winter groundwater recharge and runoff. Overall, these various possible trajectories underscore the key role of climate-vegetation feedbacks in modulating how hydroclimatic changes actually affect water balance. Given ongoing hydroclimatic shifts, process understanding of these interactions will become increasingly important to detailed projections of water flux partitioning.
4.3 Scope of the study
By focusing on a pair of instrumented sites in a long-term experimental catchment, our study design allows the key processes to be examined in detail at the appropriate scale. Nevertheless, the landscape position of the study system gives it particular significance to projections of future ecological and hydrological dynamics across the region. Through both locally higher inputs and intra-annual storage, forested montane headwater catchments play an outsize role in baseflow generation, supporting regional hydrological stability (Viviroli et al., 2007; Immerzeel et al., 2020). The broader landscape's (i.e., downstream) water regimes will be particularly sensitive to their seasonal functioning under climate change. Furthermore, the observation of an annual-scale switch from energy- to a water-limitation in a montane forest is strongly indicative for large parts of the generally warmer, drier Central European landscape.
On the other hand, the resulting process understanding is only transferable to an extent circumscribed by an adequate consideration of the landscape position of the study system. For example, while summer season temperature did not greatly affect the water balance in our study catchment, this may in part be due to comparatively low a vapour pressure deficit (VPD) at this elevation. As VPD is a strongly nonlinear function of air temperature (Grossiord et al., 2020), lower elevation forests will face disproportionately higher summer VPD, potentially increasing its importance in their water balance. This factor may also increase in importance disproportionately across the landscape with further climate warming. Given our findings, we would again expect any increased effects to be stronger in beech rather than spruce stands, due to their relatively anisohydric transpiration, and to shift the state of these systems further towards water limitation.
It should urgently be evaluated how widespread the observed deviation from the predicted trend in the seasonal timing of precipitation is. If it is merely a strong local anomaly, we would expect drier summers to exacerbate overall water-limitation and the importance of winter recharge from deciduous forest to increase over time. If the trend we found is real but limited to catchments with the specific landscape position of ours (e.g., similar exposure and position within the Bohemian Forest), then these catchments may play an offsetting role in the shifting regional water balance, smoothing out shifts in recharge. If, on the other hand, this deviation is due to a general (e.g., orographic) effect not accounted for in climate models, it may generalise to the entire Bohemian Forest and reverse expectations about both the seasonality of future water availability and, through interactions with vegetation, its annual sums in the region.
4.4 Measurement limitations
As the measuring limit of the tensiometers is −865 cm (−85 kPa), pressure heads below this limit could not be recorded. Some information was therefore lost, especially at the beech site where periods with a constant limit value were clearly visible. However, for pressure heads lower than the measurement limit, the loss and gain of the volumetric water content corresponding to the unit change in the pressure head is very small (a 100 cm change in the pressure head accounts for less than 0.002 cm3 cm−3 of the change in the volumetric water content). The same rate was observed for a saturation to a pressure head of −100 cm, which is equal to 0.22 cm3 cm−3. Hence, the changes in pressure head concerning such low heads have a negligible effect on the volumetric soil water content.
To encompass the influence of soil moisture spatial variability, 2 to 5 tensiometers were used at each depth. As the standard errors of precipitation measurements are 10 % in summer and 40 % in winter, it can be assumed that these measurements of precipitation can also be biased due to wind eddies around the rain gauge and deposited precipitation (Dingman, 2015). Even though the study sites are located close to the rain gauges (<500 m) and we also checked the open area rainfall data with the raingauges located in the forest, there were occasional episodes in the data where the volumetric water content did not match to the volume measured rainfall, which resulted in a few errors in the soil moisture modelling, especially of the rises in the volumetric soil moisture content.
4.5 Modelling limitations
Observations from the above-mentioned periods when soil pressure heads were at or below the measuring range of the tensiometer were not used to constrain or evaluate the soil water balance model. The model was allowed to run freely below this limit, and the error statistics from these periods were not considered. Eliminating this bias did not allow model fitting during dry periods. Another issue arose from the noted episodes of rainfall over- and underestimation. As both issues affected periods with negligible water fluxes, neither was found to affect the long-term water balance. Finally, as shown in Cejpek et al. (2018) and Jačka et al. (2021), different vegetation species growing on the same soil type tend to change soil properties, whether due to different root systems, soil biology or litter. Even though the soil parameters (Ks, ) that were entered into the balance model have measured equivalents at each site, their values in this study are the result of model calibration.
As we used a simple temperature-based approach for the estimation of PET we compared these estimates with state-of-the-art method of Penman-Monteith (Monteith, 1965) over the period of available data (from 2008). The influence of method selection on the resulting water fluxes was negligible (<2 % on seasonal and annual PET, AET, modelled soil water content) (see Fig. S5). The sensitivity of PET to canopy-specific aerodynamic resistance parameterisation (beech vs. spruce) in the Penman-Montieth approach was in our case outweighed by the influence of soil water availability (reflected in stomatal resistance). We limited our inferences to seasonal and annual comparisons, at which scales the differences between the PET estimation methods are negligible.
Since the model validation was performed on the average annual discharge value measured for the entire watershed, which is mostly covered by spruce forest, it is possible that these values may not correspond with the discharge that might occur from the beech site alone. This might affect confidence in the balance components (drainage and actual evapotranspiration) at the beech site as compared to the spruce site. However, the modelled high transpiration rates at the beech sites mostly follow from fitting to the high-resolution time series of measured local soil moisture data, which show lower values during the summer season compared to spruce, and simultaneous observations of no change in groundwater levels. The higher modelled transpiration rates of beech during the summer season presented in this study could also be supported by the higher measured sap flow during the summer season in Switzerland (Brinkmann et al., 2016) or nearby Kranzberg forest in Bavaria (Gebhardt et al., 2023). Moreover, due to the absence of measured soil moisture data below the tensiometer measurement limit, it could be assumed that as soil moisture values could be even lower at beech sites, transpiration will be higher than estimated. To avoid such uncertainties in future research, detailed sap flow measurements might serve for model calibration, which could then show the values of actual evapotranspiration and drainage more precisely.
Ongoing climate change is forcing a transition from energy- to water-limitation and altering the species composition of European forests. We analysed a multi-decade record of soil water potential and climatic data to determine which variables have driven water limitation so far and which vegetation processes most exacerbate or dampen it. We found evidence of annual-scale water limitation, unprecedented in Central European montane forest. While increasing atmospheric demand drives progressive water limitation at the broader scale, seasonal water supply interacts with vegetation processes to determine the actual soil water balance in the studied beech and spruce stands. Decreased summer precipitation drove stronger drying in the beech stand compared to spruce. Our water-balance model suggests that beech did not reduce transpiration rates in dry summers but continued to exploit deeper soil water reserves more extensively (by ∼100 mm per season on average), resulting in decreased drainage. During wet summers and all winter seasons, the soil was drier in the spruce stand, due to higher winter interception by its evergreen canopy (by ∼40 mm per season on average). Hence, in wet periods, drainage remained higher in the beech forest.
The results suggest that with progressing water-limitation, soil water will increasingly be disproportionately depleted in by forests composed of deeper-rooted, more anisohydric species. The combined effects of climate and forest composition change may thus increase the severity of summer soil drought and limit groundwater recharge. On the other hand, increasing the proportion of deciduous species should result in increased winter recharge, due to decreased interception by leafless canopies. As climate-vegetation interactions represent key sources of uncertainty in predicting shifts in ecosystem function and composition under climate change, we expect such advances in process understanding will contribute to the next generation of models and projections, facilitating both ecosystem and water management during the ongoing hydroclimatic shift.
The code is available from the corresponding author upon request.
The modelling data can be accessed through the public repository of the Czech Academy of Sciences (https://doi.org/10.57680/asep.0640388, Tesař et al., 2025).
The supplement related to this article is available online at https://doi.org/10.5194/hess-29-6003-2025-supplement.
VŠ, MT, LV created the concept and set the methodology; NZ, VŠ, JT, JK carried the investigation; NZ implemented the model and created visualizations; NZ, VŠ and MB wrote the manuscript draft; all authors reviewed and edited the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
The authors would like to thank David Pěsta for conducting regular field measurements and the programme framework of the Strategy AV21: Water for Life.
This research has been supported by the Grantová Agentura České Republiky (grant no. GA CR 24-10375S) and the Akademie Věd České Republiky (RVO: 67985874 and 67985939).
This paper was edited by Miriam Coenders-Gerrits and reviewed by Ronald Queck and one anonymous referee.
Arnold, J. G., Kiniry, J. R., Srinivasan, R., Williams, J. R., Haney, E. B., amd Neitsch, S. L.: Soil and Water Assessment Tool Input/Output Documentation: Version 2012, Texas Water Resources Institute, College Station, https://swat.tamu.edu/docs/ (last access: 30 October 2025), 2012.
Braden, H.: Ein Energiehaushalts-und Verdunstungsmodell for Wasser und Stoffhaushaltsuntersuchungen landwirtschaftlich genutzer Einzugsgebiete, Mittelungen Deutsche Bodenkundliche Geselschaft, 42, 294–299, 1985.
Brázdil, R., Zahradníček, P., Dobrovolný, P., Štěpánek, P., and Trnka, M.: Observed changes in precipitation during recent warming: The Czech Republic, 1961–2019, Int. J. Climatol., 41, 3881–3902, https://doi.org/10.1002/joc.7048, 2021.
Brázdil, R., Dobrovolný, P., Mikšovský, J., Pišoft, P., Trnka, M., Možný, M., and Balek, J.: Documentary-based climate reconstructions in the Czech Lands 1501–2020 CE and their European context, Clim. Past, 18, 935–959, https://doi.org/10.5194/cp-18-935-2022, 2022.
Brinkmann, N., Eugster, W., Zweifel, R., Buchmann, N., and Kahmen, A.: Temperate tree species show identical response in tree water deficit but different sensitivities in sap flow to summer soil drying, Tree Physiol., 36, 1508–1519, https://doi.org/10.1093/treephys/tpw062, 2016.
Brocca, L., Melone, F., and Moramarco, T.: On the estimation of antecedent wetness conditions in rainfall-runoff modelling, Hydrol. Process., 2274, 2267–2274, https://doi.org/10.1002/hyp.6629, 2008.
Brocca, L., Camici, S., Melone, F., Moramarco, T., Martínez-Fernández, J., Didon-Lescot, J. F., and Morbidelli, R.: Improving the representation of soil moisture by using a semi-analytical infiltration model, Hydrol. Process., 28, 2103–2115, https://doi.org/10.1002/hyp.9766, 2014.
Burgess, S. S. O., Adams, M. A., Turner, N. C., and Ong, C. K.: The redistribution of soil water by tree root systems, Oecologia, 115, 306–311, https://doi.org/10.1007/s004420050521, 1998.
Cejpek, J., Kuráž, V., Vindušková, O., and Frouz, J.: Water regime of reclaimed and unreclaimed post-mining sites, Ecohydrology, 11, 1–9, https://doi.org/10.1002/eco.1911, 2018.
Čermák, J., Cienciala, E., Kučera, J., Lindroth, A., and Bednářová, E.: Individual variation of sap-flow rate in large pine and spruce trees and stand transpiration: a pilot study at the central NOPEX site, J. Hydrol., 168, 17–27, https://doi.org/10.1016/0022-1694(94)02657-W, 1995.
Daněk, P., Šmaonil, P., and Vrška, T.: Four decades of the coexistence of beech and spruce in a Central European old-growth forest, Which succeeds on what soils and why?, Plant Soil, 437, 254–272, https://doi.org/10.1007/s11104-019-03968-4, 2019.
Denissen, J. M. C., Teuling, A. J., Pitman, A. J., Koirala, S., Migliavacca, M., Reichstein, M., Winkler, A. J., Zhan, C., and Orth, R.: Widespread shift from ecosystem energy to water limitation with climate change, Nature Climate Change, 12, 677–684, https://doi.org/10.1038/s41558-022-01403-8, 2022.
Dingman, S. L.: Physical hydrology, 3rd edn., Waveland Press, Inc., Long Grove, https://doi.org/10.1177/030913338901300106, 2015.
Feddes, R. A. and Rijtema, P. E.: Water withdrawal by plant roots, J. Hydrol., 17, 33–59, https://doi.org/10.1016/0022-1694(72)90065-0, 1972.
Floriancic, M. G., Allen, S. T., Meier, R., Truniger, L., Kirchner, J. W., and Molnar, P.: Potential for significant precipitation cycling by forest-floor litter and deadwood, Ecohydrology, 16, 1–16, https://doi.org/10.1002/eco.2493, 2022.
Gebauer, T., Horna, V., and Leuschner, C.: Canopy transpiration of pure and mixed forest stands with variable abundance of European beech, J. Hydrol., 442–443, 2–14, https://doi.org/10.1016/j.jhydrol.2012.03.009, 2012.
Gebhardt, T., Hesse, B. D., Hikino, K., Kolovrat, K., Hafner, B. D., Grams, T. E. E., and Häberle, K. H.: Repeated summer drought changes the radial xylem sap flow profile in mature Norway spruce but not in European beech, Agric. For. Meteorol., 329, 109285, https://doi.org/10.1016/j.agrformet.2022.109285, 2023.
Gerrits, A. M. J., Pfister, L., and Savenije, H. H. G.: Spatial and temporal variability of canopy and forest floor interception in a beech forest, Hydrol. Process., 24, 3011–3025, https://doi.org/10.1002/hyp.7712, 2010.
Girons Lopez, M., Vis, M. J. P., Jenicek, M., Griessinger, N., and Seibert, J.: Assessing the degree of detail of temperature-based snow routines for runoff modelling in mountainous areas in central Europe, Hydrol. Earth Syst. Sci., 24, 4441–4461, https://doi.org/10.5194/hess-24-4441-2020, 2020.
Green, J. K., Seneviratne, S. I., Berg, Am. M., Findell, K. L., Hagemann, S., Lawrence, D. M., and Gentine, P.: Large influence of soil moisture on long-term terrestrial carbon uptake, Nature, 565, 476–479, https://doi.org/10.1038/s41586-018-0848-x, 2019.
Grossiord, C., Buckley, T. N., Cernusak, L. A., Novick, K. A., Poulter, B., Siegwolf, R. T. W., Sperry, J. S., and McDowell, N. G.: Plant responses to rising vapor pressure deficit, New Phytol., 226, 1550–1556, https://doi.org/10.1111/nph.16485, 2020.
Gupta, R. S.: Hydrology and hydraulic systems, Waveland Press, Long Grove, ISBN 978-157766-030-9, 2001.
Hari, V., Rakovec, O., Markonis, Y., Hanel, M., and Kumar, R.: Increased future occurrences of the exceptional 2018–2019 Central European drought under global warming, Sci. Rep.-UK, 10, 12207, https://doi.org/10.1038/s41598-020-68872-9, 2020.
Hochberg, U., Rockwell, F. E., Holbrook, N. M., and Cochard, H.: Iso/Anisohydry: A Plant–Environment Interaction Rather Than a Simple Hydraulic Trait, Trends Plant Sci., 23, 112–120, https://doi.org/10.1016/j.tplants.2017.11.002, 2018.
Hrkal, Z., Milický, M., and Tesař, M.: Climate change in central Europe and the sensitivity of the hard rock aquifer in the Bohemian Massif to decline of recharge: Case study from the Bohemian Massif, Environ. Earth Sci., 59, 703–713, https://doi.org/10.1007/s12665-009-0067-8, 2009.
Huang, X., Shi, Z. H., Zhu, H. D., Zhang, H. Y., Ai, L., and Yin, W.: Soil moisture dynamics within soil profiles and associated environmental controls, Catena, 136, 189–196, https://doi.org/10.1016/J.Catena.2015.01.014, 2016.
Humphrey, V., Berg, A., Ciais, P., Gentine, P., Jung, M., Reichstein, M., Seneviratne, S. I., and Frankenberg, C.: Soil-moisture–atmosphere feedback dominates land carbon uptake variability, Nature, 592, 65–69, https://doi.org/10.1038/s41586-021-03325-5, 2021.
Huntingford, C., Marsh, T., Scaife, A., Kendon, E. J., Hannaford, J., Kay, A. L., Lockwood, M., Prudhomme, C., Reynard, N. S., Parry, S., Lowe, J. A., Screen, J. A., Ward, H. C., Roberts, M., Stott, P. A., Bell, V. A., Bailey, M., Jenkins, A., Legg, T., Otto, F. E. L., Massey, N., Schaller, N., Slingo, J., and Allen, M. R.: Potential influences on the United Kingdom's floods of winter 2013/14, Nat. Clim. Change, 4, 769–777, https://doi.org/10.1038/nclimate2314, 2014.
Immerzeel, W. W., Lutz, A. F., Andrade, M., Bahl, A., Biemans, H., Bolch, T., Hyde, S., Brumby, S., Davies, B. J., Elmore, A. C., Emmer, A., Feng, M., Fernández, A., Haritashya, U., Kargel, J. S., Koppes, M., Kraaijenbrink, P. D. A., Kulkarni, A. V., Mayewski, P. A., Nepal, S., Pacheco, P., Painter, T. H., Pellicciotti, F., Rajaram, H., Rupper, S., Sinisalo, A., Shrestha, A. B., Viviroli, D., Wada, Y., Xiao, C., Yao, T., and Baillie, J. E. M.: Importance and vulnerability of the world's water towers, Nature, 577, 364–369, https://doi.org/10.1038/s41586-019-1822-y, 2020.
IUSS Working Group WRB: World Reference Base for Soil Resources 2014, Update 2015, International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106, FAO, Rome, ISBN 978-92-5-108369-7, 2015.
Jačka, L., Walmsley, A., Kovář, M., and Frouz, J.: Effects of different tree species on infiltration and preferential flow in soils developing at a clayey spoil heap, Geoderma, 403, 115372, https://doi.org/10.1016/j.geoderma.2021.115372, 2021.
Jost, G., Schume, H., Hager, H., Markart, G., and Kohl, B.: A hillslope scale comparison of tree species influence on soil moisture dynamics and runoff processes during intense rainfall, J. Hydrol., 420–421, 112–124, https://doi.org/10.1016/j.jhydrol.2011.11.057, 2012.
Kofroňová, J., Šípek, V., Hnilica, J., Vlček, L., and Tesař, M.: Canopy interception estimates in a Norway spruce forest and their importance for hydrological modelling, Hydrol. Sci. J., 66, 1233–1247, https://doi.org/10.1080/02626667.2021.1922691, 2021.
Korres, W., Reichenau, T. G., Fiener, P., Koyama, C. N., Bogena, H. R., Cornelissen, T., Baatz, R., Herbst, M., Diekkrüger, B., Vereecken, H., and Schneider, K.: Spatio-temporal soil moisture patterns – A meta-analysis using plot to catchment scale data, J. Hydrol., 520, 326–341, https://doi.org/10.1016/j.jhydrol.2014.11.042, 2015.
Kuželková, M., Jačka, L., Kovář, M., Hradilek, V., and Máca, P.: Tree trait-mediated differences in soil moisture regimes: a comparative study of beech, spruce, and larch in a drought-prone area of Central Europe, Eur. J. Forest Res., 143, 319–332, https://doi.org/10.1007/s10342-023-01628-y, 2024.
Kyselý, J., Gaál, L., Beranová, R., and Plavcová, E.: Climate change scenarios of precipitation extremes in Central Europe from ENSEMBLES regional climate models, Theor. Appl. Climatol., 104, 529–542, https://doi.org/10.1007/s00704-010-0362-z, 2011.
Legates, D. R., Mahmood, R., Levia, D. F., DeLiberty, T. L., Quiring, S. M., Houser, C., and Nelson, F. E.: Soil moisture: A central and unifying theme in physical geography, Prog. Phys. Geogr., 35, 65–86, https://doi.org/10.1177/0309133310386514, 2011.
Liang, X., Lettenmaier, D. P., Wood, E. F., and Burges, S. J.: A simple hydrologically based model of land surface water and energy fluxes for general circulation model, J. Geophys. Res., 99, 2063, https://doi.org/10.1143/jjap.39.2063, 1994.
Maxwell, T. M., Silva, L. C. R., and Horwath, W . R.: Integrating effects of species composition and soil properties to predict shifts in montane forest carbon–water relations, P. Natl. Acad. Sci. USA, 115, E4219–E4226, https://doi.org/10.1073/pnas.1718864115, 2018.
Mianabadi, A., Davary, K., Pourreza-Bilondi, M., and Coenders-Gerrits, A. M. J.: Budyko framework; towards non-steady state conditions, J. Hydrol., 588, 125089, https://doi.org/10.1016/j.jhydrol.2020.125089, 2020.
Milly, P. C. D., Betancourt, J., Falkenmark, M., Hirsch, R. M., Kundzewicz, Z. W., Lettenmaier, D. P., Stouffer, R. J., Dettinger, M. D., and Krysanova, V.: On critiques of “Stationarity is dead: Whither water management?”, Water Resour. Res., 51, 7785–7789, https://doi.org/10.1002/2015WR017408, 2015.
Monteith, J. L.: Evaporation and environment, Symposia of the Society for Experimental Biology, 19, 205–234, 1965.
Možný, M., Trnka, M., Vlach, V., Vizina, A., Potopová, V., Zahradníček, P., Štěpánek, P., Hajková, L., Staponites, L., and Žalud, Z.: Past (1971–2018) and future (2021–2100) pan evaporation rates in the Czech Republic, J. Hydrol., 590, 125390, https://doi.org/10.1016/j.jhydrol.2020.125390, 2020.
Novick, K. A., Ficklin, D. L., Baldocchi, D., Davis, K. J., Ghezzehei, T. A, Konings, A. G., MacBean, N., Raoult, N., Scott, R. L., Shi, Y., Sulman, B. N., and Wood, J. D.: Confronting the water potential information gap, Nat. Geosci., 15, 158–164, https://doi.org/10.1038/s41561-022-00909-2, 2022.
Oudin, L., Hervieu, F., Michel, C., Perrin, C., Andréassian, V., Anctil, F., and Loumagne, C.: Which potential evapotranspiration input for a lumped rainfall-runoff model? Part 2 – Towards a simple and efficient potential evapotranspiration model for rainfall-runoff modelling, J. Hydrol., 303, 290–306, https://doi.org/10.1016/j.jhydrol.2004.08.026, 2005.
Qing, Y., Ancell, B. C., and Yang, Z.-L.: Accelerating flash droughts induced by the joint influence of soil moisture depletion and atmospheric aridity, Nat. Commun., 13, 1139, https://doi.org/10.1038/s41467-022-28752-4, 2022.
Renner, M., Brust, K., Schwärzel, K., Volk, M., and Bernhofer, C.: Separating the effects of changes in land cover and climate: a hydro-meteorological analysis of the past 60 yr in Saxony, Germany, Hydrol. Earth Syst. Sci., 18, 389–405, https://doi.org/10.5194/hess-18-389-2014, 2014.
Rötzer, T., Häberle, K. H., Kallenbach, C., Matyssek, R., Schütze, G., and Pretzsch, H.: Tree species and size drive water consumption of beech/spruce forests – a simulation study highlighting growth under water limitation, Plant Soil, 418, 337–356, https://doi.org/10.1007/s11104-017-3306-x, 2017.
Savenije, H. H. G.: The importance of interception and why we should delete the term evapotranspiration from our vocabulary, Hydrol. Process., 18, 1507–1511, https://doi.org/10.1002/hyp.5563, 2004.
Schafstall, N., Svitavská-Svobodová, H., Kadlec, M., Gałka, M., Kuneš, P., Bobek, P., Goliáš, V., Pech, P., Nývlt, D., Hubený, P., Kuosmanen, N., Carter, V. A., and Florescu, F.: The absence of disturbances promoted Late Holocene expansion of silver fir (Abies alba) in the Bohemian Forest, Palaeogeogr. Palaeoclimatol. Palaeoecol., 635, 111950, https://doi.org/10.1016/j.palaeo.2023.111950, 2024.
Schume, H., Jost, G., and Hager, H.: Soil water depletion and recharge patterns in mixed and pure forest stands of European beech and Norway spruce, J. Hydrol., 289, 258–274, https://doi.org/10.1016/j.jhydrol.2003.11.036, 2004.
Schwärzel, K., Menzer, A., Clausnitzer, F., Spank, U., Häntzschel, J., Grünwald, T., Köstner, B., Bernhofer, C., and Feger, K. H.: Soil water content measurements deliver reliable estimates of water fluxes: A comparative study in a beech and a spruce stand in the Tharandt forest (Saxony, Germany), Agric. For. Meteorol., 149, 1994–2006, https://doi.org/10.1016/j.agrformet.2009.07.006, 2009.
Seibert, J. and Vis, M. J. P.: Teaching hydrological modeling with a user-friendly catchment-runoff-model software package, Hydrol. Earth Syst. Sci., 16, 3315–3325, https://doi.org/10.5194/hess-16-3315-2012, 2012.
Šípek, V. and Tesař, M.: Seasonal snow accumulation in the mid-latitude forested catchment, Biologia, 69, 1562–1569, https://doi.org/10.2478/s11756-014-0468-3, 2014.
Šípek, V. and Tesař, M.: Year-round estimation of soil moisture content using temporally variable soil hydraulic parameters, Hydrol. Process., 31, 1438–1452, https://doi.org/10.1002/hyp.11121, 2017.
Šípek, V., Hnilica, J., Vlček, L., Hnilicová, S., and Tesař, M.: Influence of vegetation type and soil properties on soil water dynamics in the Šumava Mountains (Southern Bohemia), J. Hydrol., 582, 124285, https://doi.org/10.1016/j.jhydrol.2019.124285, 2020.
Svoboda, V., Hanel, M., Máca, P., and Kyselý, J.: Characteristics of rainfall events in regional climate model simulations for the Czech Republic, Hydrol. Earth Syst. Sci., 21, 963–980, https://doi.org/10.5194/hess-21-963-2017, 2017.
Tesař, M., Šípek, V., Zelíková, N., Toušková, J., and Vlček, L.: Hydrometeorological data from the Liz basin, Knihovna Akademie Věd ČR [data set], https://doi.org/10.57680/asep.0640388, 2025.
Tolasz, R., Míková, T., Valeriánová, A., et al.: Climate Atlas of Czechia, Czech Hydrometeorological Institute, Prague, ISBN 978-80-86690-26-1, 2007.
Toušková, J., Falátková, K., and Šípek, V.: Estimating potential evapotranspiration in a temperate zone: The challenge of model selection, Water Res. Manag., 39, 5911–5927, https://doi.org/10.1007/s11269-025-04233-3, 2025.
Trugman, A. T., Medvigy, D., Mankin, J. S., and Andregg W. R. L.: Soil moisture as a major driver of carbon cycle uncertainty, Geophys. Res. Lett., 45, 6495–6503, https://doi.org/10.1029/2018GL078131, 2018.
Tyree, M. T. and Zimmermann, M. H.: Xylem structure and the Ascent of Sap, Springer, Heidelberg, https://doi.org/10.1007/978-3-662-04931-0, 2002.
Van Genuchten, M. T.: A closed-form equation for predicting the hydraulic conductivity of unsaturated soils, Soil Sci. Soc. Am. J., 44, 892–898, https://doi.org/10.2136/sssaj1980.03615995004400050002x, 1980.
Viviroli, D., Dürr, H. H., Messerli, B., Meybeck, M., and Weingarten, R.: Mountains of the world, water towers for humanity: Typology, mapping, and global significance. Water Resour. Res., 43, 1–13, https://doi.org/10.1029/2006WR005653, 2007.
Vondráková, A., Vávra, A., and Voženílek, V.: Climatic regions of the Czech Republic, J. Maps, 9, 425–430, https://doi.org/10.1080/17445647.2013.800827, 2013.
von-Hoyningen-Huene, J.: Die interzeption des Niederschlages in landwirtschaftlichen Pflanzenbeständen, DVWK, 57, 3–51, 1983.
Wang, H., Tetzlaff, D., and Soulsby, C.: Modelling the effects of land cover and climate change on soil water partitioning in a boreal headwater catchment, J. Hydrol., 558, 520–531, https://doi.org/10.1016/j.jhydrol.2018.02.002, 2018.
Yue, S., Pilon, P., Phinney, B., and Cavadias, G.: The influence of autocorrelation on the ability to detect trend in hydrological series, Hydrol. Process., 16, 1807–1829, https://doi.org/10.1002/hyp.1095, 2002.
Zahradníček, P., Brázdil, R., Štěpánek, P., and Trnka, M.: Reflections of global warming in trends of temperature characteristics in the Czech Republic, 1961–2019, Int. J. Climatol., 41, 1211–1229, https://doi.org/10.1002/joc.6791, 2020.
Zucco, G., Brocca, L., Moramarco, T., and Morbidelli, R.: Influence of land use on soil moisture spatial-temporal variability and monitoring, J. Hydrol., 516, 193–199, https://doi.org/10.1016/j.jhydrol.2014.01.043, 2014.
Zweifel, R., Böhm, J. P., and Häsler, R.: Midday stomatal closure in Norway spruce – Reactions in the upper and lower crown, Tree Physiol., 22, 1125–1136, https://doi.org/10.1093/treephys/22.15-16.1125, 2002.