the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 24 Nov 2025
| 24 Nov 2025
Sub-daily stable water isotope dynamics of urban tree xylem water and ambient vapor
Ann-Marie Ring
Dörthe Tetzlaff
Christian Birkel
Chris Soulsby
The sub-daily dynamics of vegetation mediated water cycling and how this changes over the growing season remains largely unexplored due to measurement constraints. The monitoring of long-term in situ high-resolution stable water isotopes can provide valuable insights into how trees and the surrounding atmosphere cycle water under different conditions. In this study, we analyzed a sub-daily (∼3–4 hourly) dataset of atmospheric water vapor (δv) and tree stem xylem water (δxyl) in an urban tree stand in Berlin, Germany. We compared the diurnal (24 h) patterns of water cycling in δv and δxyl values, as well as ecohydrological variables during a summer drought (1 July–14 August) followed by a rewetting period (15 August–30 September) in 2022. Over the summer drought, water cycling was predominantly radiation driven, with highest vapor pressure deficit (VPD) rates in the afternoons and persistently dry soils. We found systematic behaviour in both δv and δxyl signatures during the summer drought and δv values were characterized by a daytime depletion in heavy isotopes, driven by local evaporation and atmospheric factors (i.e., entrainment). The mean amplitude between day- and night-time values of δv during drought was 26 ‰ for δ2H. Daytime enrichment in δxyl values, with maximal isotopic enrichment in afternoons, was consistent with diurnal hydroclimatic cycles, maximum potential evaporation (PET) of ∼ 0.75 mm h−1 limited sap flow sourced from enriched soil water at the topsoil and stomatal regulation of transpiration. The mean amplitude between day- and night-time values of δxyl during drought was 38 ‰ for δ2H. The trees showed lower twig water potential and sap flux relative to VPD in the afternoons, as well as stagnated night-time stem swelling, but the mature trees could overall sustain their physiological functioning. During rewetting, the water cycle of the green space was precipitation driven, while maximum PET rates decreased to 0.5 mm h−1. The systematic diurnal cycling of δv values was mostly discontinued due to lower soil and canopy evaporation. Only δv values just above the grassland surface (0.15 m) showed a significant daytime enrichment in heavy isotopes (amplitude ∼36 ‰ δ2H) hinting for evapotranspiration fluxes promoted by high moisture stored in soil and vegetation surfaces and transpiration of superficially enriched soil water. δxyl values were still characterized by significant daytime enrichment, however, with sub-daily amplitudes of 20 ‰ δ2H almost halved compared to the drought period, when hydraulic conductance was restricted. Our continuous, sub-daily dataset of δv and δxyl values provides unique insights on the complex stable water isotope dynamics within trees and the surrounding atmosphere. It has the potential to help constrain ecohydrological models towards prediction of climate change impacts on water cycling within vegetated areas.
- Article
(6601 KB) - Full-text XML
-
Supplement
(3276 KB) - BibTeX
- EndNote




Terrestrial vegetation plays a critical role in regulating water and energy fluxes (Asbjornsen et al., 2011; Bernacchi and VanLoocke, 2015). These fluxes are driven by the ecohydrological dynamics of atmospheric vapor and tree-internal water transport, which are essential for assessing evapotranspiration (ET) patterns, water-limiting conditions, tree function, and groundwater recharge (Dubbert and Werner, 2019; Stevenson et al., 2025; Wang et al., 2017). Despite their importance, these water cycling processes have rarely been investigated on sub-daily scales over prolonged periods mainly due to measurement constraints (Warix et al., 2023; Penna et al., 2018; Meili et al., 2021). Differentiating between morning, afternoon and nighttime temporal patterns, including source water partitioning of water fluxes such as hydraulic redistribution, transpiration cycling and relative humidity gradients brings a novel perspective to ecohydrology (Nadezhdina et al., 2010; Konarska et al., 2016; Kim et al., 2021; Stevenson et al., 2023). Seasonal changes, along with alternating wet and dry periods (DWD, 2022a), are of additional importance for a holistic understanding of water regimes in vegetated landscapes. In particular extreme events like summer droughts and subsequent autumn rewetting remain poorly understood at sub-daily scales, limiting insight into tree water use (Marx et al., 2022). As a result, how these sub-daily dynamics evolve over the growing season and affect water cycling has yet to be fully elucidated to predict further impact of climate variability on vegetation water fluxes (Seeger and Weiler, 2021; Ring et al., 2024).
Stable water isotopes (SWI) are useful tools for understanding water cycling in the soil-plant-atmosphere continuum (SPAC) (Tetzlaff et al., 2015). Numerous studies have investigated water cycling through the lens of SWI dynamics in plant xylem water (δxyl) and atmospheric water vapor (δv) in the SPAC (e.g. Orlowski et al., 2023; Dubbert and Werner, 2019). The collection of δxyl can be performed destructively by drilling xylem cores or clipping branches in the field and most commonly extracting the water via cryogenic vacuum distillation in the lab – mostly at coarse temporal resolution from weeks to months (Tetzlaff et al., 2021) or at higher frequency but usually over a relatively short amount of days (Sohel et al., 2023a; Bernhard et al., 2024). In the past, most samples of δv were obtained through discrete collection in the field via cryogenic trapping (e.g. Helliker et al., 2002). Also, a more novel method is trapping soil water vapor with diffusion-tight bags (Dahlmann et al., 2025; Herbstritt et al., 2023).
More recently, field studies which measure SWI in situ by cavity ring-down spectroscopy (CRDS) have provided more detailed insights into processes of water cycling (Beyer et al., 2020; Galewsky et al., 2016). For instance, original observations were made of the SWI dynamics in soil and transpiration water of mixed tree cultures as well as pore water in the subsurface of a vineyard terrace (Volkmann et al., 2016a; Volkmann and Weiler, 2014). After the pioneering approach for in situ monitoring of tree δxyl values by Volkmann et al. (2016b), various in situ studies investigating δxyl at high temporal resolution were performed over several weeks or months, e.g. for beech trees in a mixed forest (Gessler et al., 2022) or riparian willow trees (Landgraf et al., 2022), and rarely at urban sites, usually presented as daily mean values to account for the high methodological uncertainties (Kühnhammer et al., 2022; Landgraf et al., 2022). Wherein, the fractionation of δxyl values through plant physiological processes can be a confounding issue (Barbeta et al., 2020; Vega-Grau et al., 2021). The influence of time of the day on in situ monitored δxyl was first investigated in a tropical field location during the dry season for three months by Kühnhammer et al. (2022). They found clear diurnal differences of δxyl; although data analysis was restricted with <1.2 measurements per day for each of around 3 boreholes of two trees and soil moisture measurements at different depths. Kübert et al. (2022) have analyzed the dynamics of sub-daily transpired water from an experiment in an enclosed rainforest using a δ2H deep water pulse. They were able to capture quick responses (2 h resolution) in plant water transport showing the value of high resolution in situ measurements of plant water. Recent work by Tierney et al. (2025) on the influence of time of day on xylem SWI samples indicated clear differences between AM and PM samples of δxyl values of oak and maple in a temperate US forest. To our knowledge, no literature exists which combined prolonged in situ measurements of δxyl and δv in combination with high-resolution sub-daily environmental and plant physiological data.
Several in situ studies that have performed real-time analysis of δv provided insights on seasonal changes in partitioning of ET above agricultural land (Wei et al., 2015) or diurnal variations in evaporative signals at different heights above the Greenland ice sheet (Steen-Larsen et al., 2013). A recent urban study from Berlin-Friedrichshagen, 2021, has shown contrasting (sub-)daily dynamics of δv above different urban green spaces, that indicated evaporative fractionation after rainfall events just above an urban grassland surface (Ring et al., 2023). But in general, prolonged (several months) in situ measurements of δxyl and δv and that allow interpretation of sub-daily dynamics over a full growing period are rare. Reasons are that obtaining and quality-checking such datasets is time consuming and expensive. Previously reported methodological uncertainties can be overcome through rigorous maintenance and quality checking (cf. methods by Ring et al., 2024; Kühnhammer et al., 2022).
Here, we analyze a sub-daily dataset of in situ δxyl and δv together with environmental and plant physiological data from an urban tree stand and an urban grassland throughout the whole growing season in 2022, in Berlin, Germany. We compare diurnal (24 h) patterns of water cycling at these different types of vegetation during a period of summer drought followed by autumn rewetting. The summer in 2022 was the 4th hottest summer in Berlin since records began in 1881, according to DWD (2022c). Thus, this research has the potential to gain novel high-frequency insights on sub-daily atmospheric vapor and plant water cycling in times of climate change.
Our specific research questions were:
-
What are the sub-daily dynamics of δxyl and δv valuesover the growing season in different types of vegetation?
-
Can we identify water limitations on tree function at a sub-daily basis?
-
Do high-resolution sub-dailywater isotope data provide significant new insights, or are daily means sufficient for understanding moisture cycling at the plot scale?
The work was intended to give valuable insights into the mechanistic links of sub-daily water cycling at the SPAC during summer drought and autumn rewetting. It was also expected that the sub-daily signals help to explain previously unexplained short-term variations in daily SWI datasets.
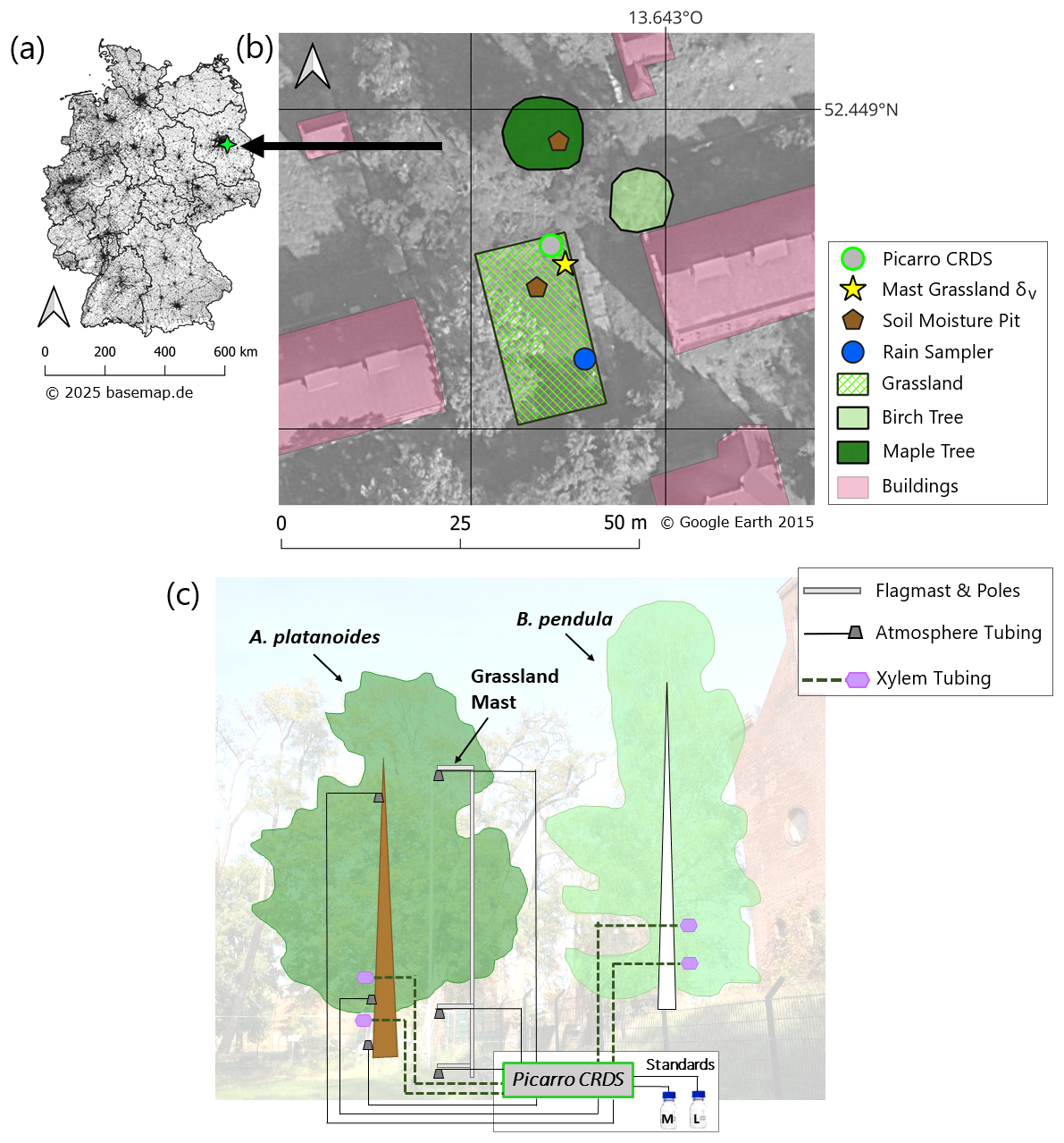
Figure 1(a) Location of the study site in Germany (residential areas and borders of federal states in black – modified from basemap.de/open-data). (b) Study site in Berlin-Friedrichshagen with investigated grassland, A. platanoides and B. pendula, including cavity ring-down spectroscopy (CRDS), flag mast for atmospheric vapor (δv) measurements, soil moisture pits (underneath canopy and at grassland) and precipitation sampler (basemap: © Google Earth 2015). (c) Conceptual diagram of the general in situ measuring setup for stable water isotopes of atmospheric water vapor (δv) and plant xylem water (δxyl). All inlets were measured sequentially via tubing connected to the CRDS. δv was measured at 0.15, 2 and 10 m height under the A. platanoides canopy and above grassland at a flag-pole (distance between A. platanoides and flag mast was 16 m; grey caps refer to rain and wind protection cover). δxyl was measured from boreholes at A. platanoides and B. pendula (1.5 and 2.5 m height; violet hexagons refer to attached bottles filled with desiccant). Temperature probes were placed at each tubing inlet. Standards were connected via tubing for automated calibration with liquid values of δ2H = −73.623 ‰ and δ18O = −10.522 ‰ for the “L” standard, and δ2H = 16.74 ‰ and δ18O = 1.53 ‰ for the “M” standard.
The study was conducted in a peri-urban area southeastof Berlin, Germany, on the grounds of the LeibnizInstitute of Freshwater Ecology and Inland Fisheries (IGB) (Fig. 1a). Berlin is situated around the former Warszaw-Berlin ice marginal valley and its glacial spillway which were formed during the Weichselian glaciation (Lüthgens and Böse, 2011). From this emerged a flat topography filled with sand and gravel deposits (Limberg et al., 2007). 59.4 % of Berlin is residential areas, followed by 34 % with vegetation cover and 6.6 % surface waters (SenMVKU, 2023). The study site is located roughly 200 m north of Lake Müggelsee, 38 m a.s.l. It is surrounded by buildings (on average 10 m distance) and vegetation is a mosaic of extensive grassland and mature trees (∼30–100 years old). Groundwater levels directly at the site are ∼4 m below the ground surface (Umweltatlas Berlin, 2020) with limited annual variability (<0.15 m). Sandy regosols are the prevailing soil type (Geoportal Berlin, 2015) and anthropogenic impacts are reflected through existing debris, sandy materials and a shallow humus layer (roughly 8 cm deep).
Berlin's climate is a warm-summer humid continental climate (Köppen classification: Dfb; Beck et al., 2018). The long-term (1991–2020) mean annual precipitation and temperature are 579 mm and 10.2 °C, respectively (DWD, 2023). In comparison to the 1981–2010 reference period, the region has become warmer and drier in recent years, making it more susceptible to drought. In 2022, the study year, Berlin only received 403 mm of annual rainfall (DWD, 2024). Summer precipitation exhibits irregular, more intensive, convective rainfall (SenStadt, 2025). This high-intensity precipitation during summer is often lost to urban drainage or high ET responses, and often does not fully replenish soil moisture or recharge groundwater.
This study focused on two different types of vegetation: a grassland and a tree-dominated area, both non-irrigated (Fig. 1b). The grassland was managed at low intensity (mowed twice a year) and covered by grass (e.g. Bromus sterilis, Lolium perenne) and herb species (e.g. Rumex acetosa, Trifolium pratense) of up to 60 cm height. For the investigation of urban trees, we focused on one large A. platanoides tree (“norway maple”; ∼80 years old) with a height of ∼16 m and a stem diameter of 560 mm and one dominant B. pendula tree (“silver birch”; ∼60 years old) with a height of ∼12 m and a stem diameter of 490 mm (stem diameters were measured in April 2022). Both tree genera are representative of temperate-zone city trees (Roloff, 2013). Previous isotope-based studies at the site have detected deep soil water and groundwater as the likely dominant sources of water to these trees (Ring et al., 2024) and an enrichment of atmospheric water vapor just above the grassland surface indicating evaporative enrichment (Ring et al., 2023).
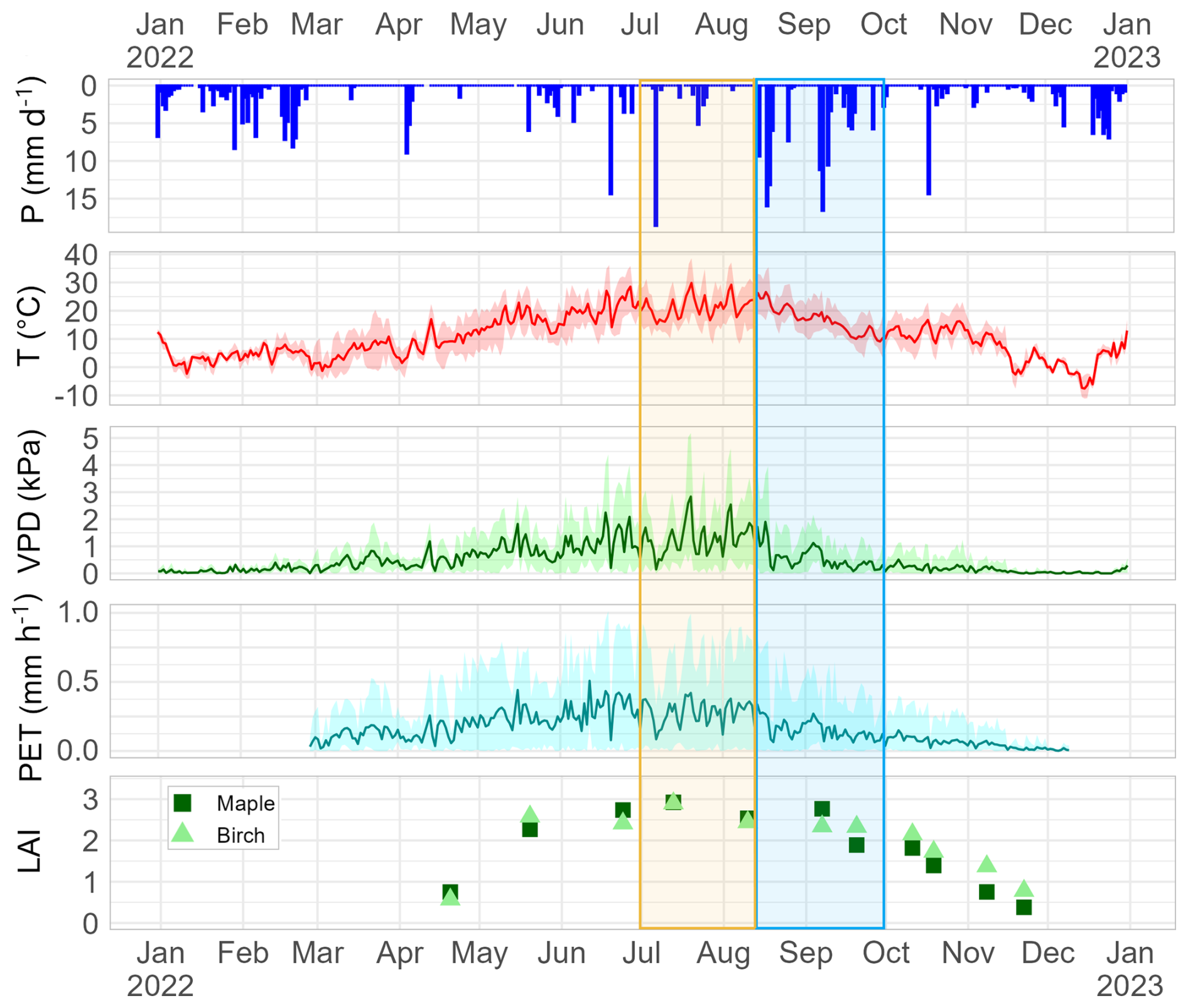
Figure 2Hydroclimatic Context 2022: Precipitation [mm d−1] is shown as daily sum. Temperature [°C], VPD [kPa] and PET [mm h−1] are shown as daily mean values with daily min/max band. Leaf Area Index (LAI) is shown as daily mean values. Periods of interest are shown as an orange box (summer drought 1 July–14 August 2022) and blue box (rewetting 15 August–30 September 2022). Figure adapted from Ring et al. (2024).
3.1 Climatic inputs and precipitation isotopes
This study focuses particularly on two sub-periods within the growing season in Berlin, 2022: a summer drought period from 1 July to 14 August and a rewetting period from 15 August to 30 September. Details of climate data are summarised in Table S1 and Fig. 2. Hourly potential evapotranspiration (PET [mm h−1]; cf. Bliss Singer et al., 2021) was estimated by the FAO Penman–Monteith method (Allen et al., 1998) using data from a IGB rooftop station 315 m away from the study site, as this featured the most continuous timeseries. The transition point between the drought and the rewetting period was identified for the 15 August 2022 with the onset of late summer rain. This choice was based on a study outlined by Birkel et al. (2025b), which combined data from an associated field study at this site with parsimonious tracer-aided conceptual modelling. Their analysis showed that both hydrometeorology and stable water isotopic signatures rapidly changed from dry to wet on that day in 2022.
Precipitation amounts, as well as precipitation and groundwater isotopes were measured at the study site and on IGB grounds (see Table S1 in the Supplement). Details on handling of liquid water samples can be found in Ring et al. (2024). Expression of results was in δ-notation with Vienna Standard Mean Ocean Water (VSMOW).
3.2 In situ monitoring of δv and δxyl
SWI of atmospheric water vapor (δv) and plant xylem water (δxyl) were measured in situ real-time and sequentially (automatic switch of tube) via a cavity ring-down spectroscopy (CRDS) laser (L2130-i, PICARRO, INC., Santa Clara, CA; Fig. 1c). The results were expressed in δ-notation as the ratio of heavy over light SWI with relation to Vienna Standard Mean Ocean Water (VSMOW). The CRDS laser was installed between the trees and the grassland in a shaded box to prevent from overheating during high radiation exposure. Measurement details are displayed in TableS1 in the Supplement. The plot-scale elevation profile above the two distinct types of vegetation was monitored to assess possible effects of diverse urban vegetation and potential air turbulence. δv was measured at 0.15, 2 and 10 m height at the urban grassland using a flag mast and under the canopy of the A. platanoides (Fig. 1c). Each δv tube inlet (Fig. 1c) was sampled for 20 min. The first 8 min after tube switching were always discarded to mitigate memory effects only data exhibiting stable isotope values were retained for analysis (defined as standard deviation of ≤2 ‰ for δ2H and ≤0.5 ‰ for δ18O). To prevent the δv inlets from rain and radiation exposure, we fitted PET plastic-bottles covered with aluminum foil over each δv tube inlet.
δxyl values of A. platanoides and the B. pendula were monitored in situ via the stem borehole equilibration method (Kühnhammer et al., 2022; Marshall et al., 2020; Beyer et al., 2020). At each tree, two stem boreholes (1.5 and 2.5 m height) were drilled to record potential height variations or travel times within the tree stems. Details on the method are given in Ring et al. (2024). Each δxyl tube inlet (Fig. 1c) was sampled for 30 min. To mitigate memory effects, the first 15 min of δxyl measurements following a tube switch were consistently excluded, and only data exhibiting stable isotope values were retained for analysis (defined as standard deviation of ≤3 ‰ for δ2H and ≤0.8 ‰ for δ18O). Changes in water concentration of the analyzed water vapour have an effect on SWI measurements with concentrations below 10 000 ppm leading to unstable SWI values (Haberstroh et al., 2024; Wassenaar et al., 2008). In the field, in-situ measurements are also subject to temperature fluctuations and especially borehole δxyl measurements should reach high water saturation to assume equilibrium conditions (min. 80 %; Marshall et al., 2020). We have checked the sub-daily water vapor concentration change for the SWI sampling inlets and calculated the saturation point for the respective temperatures (cf. ∼ 8500 ppm at 5 °C and ∼41 700 ppm at 30 °C). Water vapor concentration of the SWI data analyzed in this study was always close to full saturation during the measurements (e.g. 28 000 ppm at 25 °C which has a saturation point at ∼31 400 ppm). Thus, we can rule out evaporation effects and kinetic fractionation of measurements. The water vapor concentration range covering the whole drought and rewetting period (1 July–30 September 2022) is displayed in Fig. S11. We could not use the δxyl data of the A. platanoides borehole at 2.5 m height for further analysis due to unstable measurements, which simultaneously occurred with high CH4 values detected by the CRDS (cf. Ring et al., 2024).
3.3 Liquid δ-values and calibration
For later conversion of δv and δxyl measurements into liquid water isotope values and to account for temperature dependencies, temperature probes were set up at all tube inlets (see Table S1). By combining the in situ stable water isotope data with associated temperature data we could correct the measured values of δv and δxyl to liquid values with the correction Eq. (1) formulated by Majoube (1971):
where α is the isotopic fractionation factor, Tk is the temperature (in K), and a, b, and c are empirical parameters that vary depending on the isotopologue (values in Supplement).
Therefore, all isotopic values in this study can be compared with each other, as they are all given for the liquid phase and relative to Vienna Standard Mean Ocean Water (VSMOW). To correct for isotopic offsets and vapor concentration dependencies, automated calibration of CRDS measurements was performed after every three loops of measuring δv and δxyl values using two standards, which had liquid isotope values of δ2H = −73.623 ‰ and δ18O = −10.522 ‰ for the “light” standard, and δ2H = 16.74 ‰ and δ18O = 1.53 ‰ for the “heavy” standard. Then, temperature-dependent slopes were incorporated using linear regression.
3.4 Soil water isotopes
Further, to detect the variation in bulk soil water isotopic signatures over time and between the grassland and the tree site, we conducted monthly destructive soil sampling from April until October 2022 at the grassland site and underneath A. platanoides (Fig. 1b) at five depths (0–5, 5–10, 10–20, 20–40 and 40–70 cm) (see Table S1). For analysis in the lab, we used the direct equilibrium method outlined by Wassenaar et al. (2008). Further details on the method are displayed in Ring et al. (2024).
3.5 Ecohydrological parameters
To gain full insights into ecohydrological partitioning at our study site, we also implemented comprehensive monitoring of ecohydrological parameters, that reflect changes in water status of the soil-plant-atmosphere-continuum (SPAC): including sap flow, stem growth, twig water potential, LAI, soil moisture and groundwater levels (see Table S1). Sap flow rates were assessed via the monitored sap velocity (heat ratio method by Marshall (1958) with the softwares implexx (SFM-4 m, UGT) and Sap Flow Tool (SFM1, ICT International) including data of sap wood, heart wood and bark depths from drilled tree cores. Initially, we corrected the sap velocity raw data with a zero-flow offset. The soil moisture monitoring pits entirely consisted of sandy soils (over 94 % of fractions >0.063 mm).
3.6 Data analysis
Data processing and analysis were conducted in R, Version 4.4.1 (R Core Team, 2024). We used the Shapiro-Wilk test (Shapiro and Wilk, 1965) to test the normality of all datasets. For the non-normally distributed isotopic data we used non-parametric alternatives (assuming observations of SWI were paired and random): the Wilcoxon signed-rank test for two groups of day and night values (Wilcoxon, 1945), the Friedman test by ranks for plus two groups (Friedman, 1937) and post-hoc the pairwise Wilcoxon rank sum test with Bonferroni correction (Bonferroni, 1935; Mann and Whitney, 1947; Wilcoxon, 1945).
For the assessment of evaporation, the line-conditioned excess (short lc-excess) (Landwehr and Coplen, 2006) of the measured water isotopic signatures was calculated for local evaporative effects. Lc-excess [‰] represents the deviation of the sample from the local meteoric water line (LMWL):
where a is the slope and b the intercept of the weighted isotopic composition of the local precipitation. The LMWL was calculated by amount-weighted least square regression (Hughes and Crawford, 2012) from daily precipitation isotopes measured at the study site during the whole year of 2022, where a=7.37 and b=4.249.
To assess potential time-dependent relationships between the monitored ecohydrological variables and high-resolution in situ isotope data of δxyl and δv, we performed a cross-correlation analysis at hourly resolution. This method is commonly used in hydrology to identify connections and trends between hydrological and meteorological timeseries (Khaliq et al., 2009; Rahmani and Fattahi, 2021). It computes the relationship between two univariate (time-) series as a function of the (time-) lag between them (cf. Chambers et al., 2002). The in situ measured isotopes were aggregated to hourly data via linear interpolation for cross-correlations.
4.1 Hydroclimatic conditions during 2022
Hydroclimatic conditions surrounding the study period in 2022 are shown in Fig. 2. Compared to the international reference period 1961–1990 the spring of 2022 was very dry with very low precipitation input from March until May with ∼40 mm at study site within these 3 months compared to a long-term average of 132 mm for Berlin (DWD, 2022b). The summer 2022 was also very dry and warm in the area of Berlin with 20.6 °C and 120 mm compared to 17.8 °C and 182 mm between 1961–1991. (DWD, 2022c).
With regard to the identified summer drought period (1 July–14 August 2022) the daily mean temperature was 21.6 °C (cf. long-term average 1991–2020: 19.7 °C; DWD, 2023), reaching a maximum of 38.5 °C. Cumulative precipitation at the study site during that period was 32.4 mm (long-term average sum Berlin 1991–2020: 105 mm; DWD, 2023), with most events totaling less than 5 mm. The largest rainfall event was a convective event of 18.6 mm on 7 July. Daily maximum PET during the summer drought was mostly above 0.6 mm h−1. Daily mean VPD exceeded 1 kPa throughout most of the drought period. The identified rewetting period started from 15 August with recurrent summer rain events of >10 mm and continued with frequent rainfall in September. Daily mean PET was mostly <0.25 mm h−1 during rewetting.
LAI depicted rapid leaf-burst between April and May and showed highest values between June and early September. From late September until November the advancing leaf senescence was represented in LAI. Twig water potential of both trees (Fig. S1, Supplement) was most negative in July and August, compared to spring and autumn months. Additionally, it was more negative during midday compared to morning measurements during the drought period (Fig. S1, Supplement).
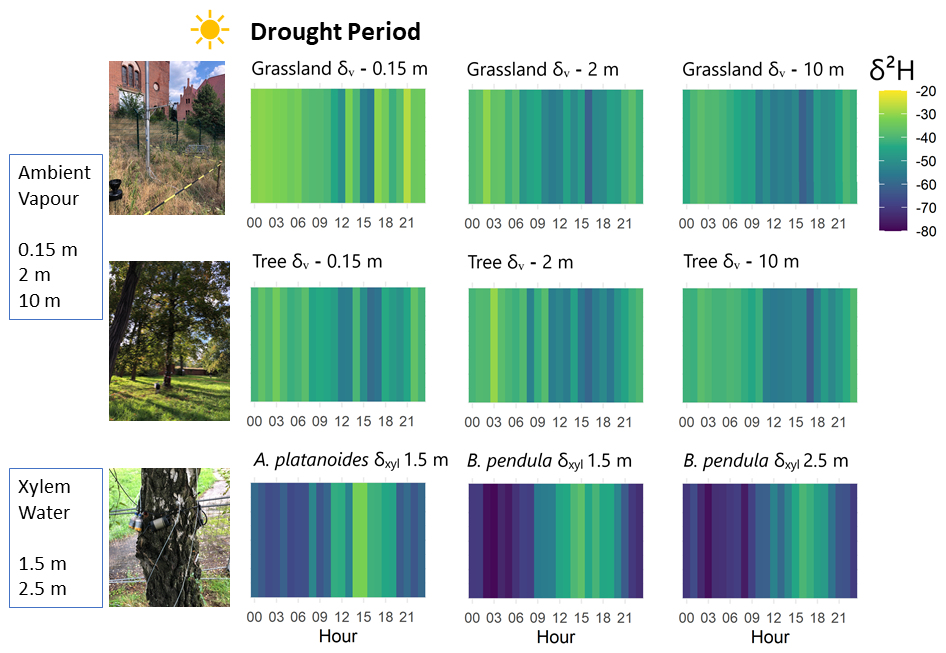
Figure 324 h cycle heatmap of hourly median δ2H data of tree xylem water (δxyl) and atmospheric vapor (δv) during summer drought period 1 July–14 August 2022. One in situ measurement loop took ∼3.5 h and unstable measurements were ruled out (cf. Sect. 3). The hourly median shown for each hour of the day was calculated based on ∼8.6 measurements.
4.2 Sub-daily Dynamics of Stable Isotopes
In this section, we focus on δ2H values, setting aside δ18O values which showed near-identical patterns (displayed in the Supplement as Figs. S2, S3 and S4). High-resolution in situ monitoring of δxyl and δv values exhibited clear diurnal cycling during the entire drought period (1 July–14 August 2022; Fig. 3). Wilcoxon-tests on differences between day (08:00 a.m.–08:00 p.m. CEST) and night (08:00 p.m.–08:00 a.m. CEST) isotopic signatures indicated significant (p<0.01) statistical difference for δxyl and δv values. However, opposing systematic sub-daily dynamics in the isotope signatures of ambient vapor and plant xylem water were apparent: δv values showed afternoon depletion and nighttime enrichment. δxyl values showed clear afternoon enrichment in heavy isotopes in both trees indicating circadian cycle. Especially, δxyl values of the B. pendula became highly depleted (up to −80 ‰ δ2H) during nighttime. The mean amplitude between day- and night-time values of δxyl, averaged over all trees, was 38 ‰ δ2H (Fig. 6).
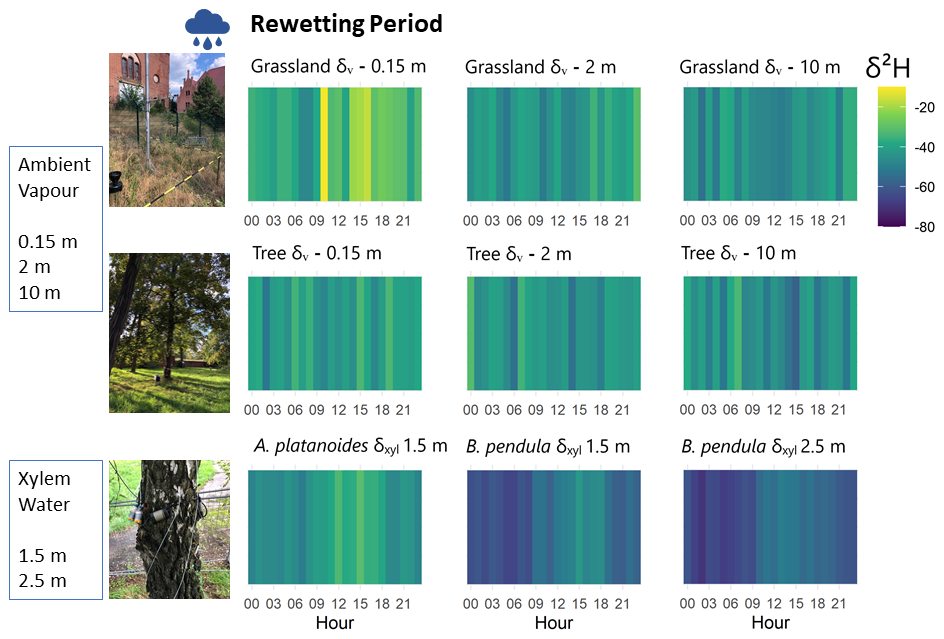
Figure 424 h cycle heatmap of hourly median δ2H data of tree xylem water (δxyl) and atmospheric vapor (δv) during rewetting period 15 August–30 September 2022. One in situ measurement loop took ∼3.5 h and unstable measurements were ruled out (cf. Sect. 3). The hourly median shown for each hour of the day was calculated based on ∼9.2 measurements).
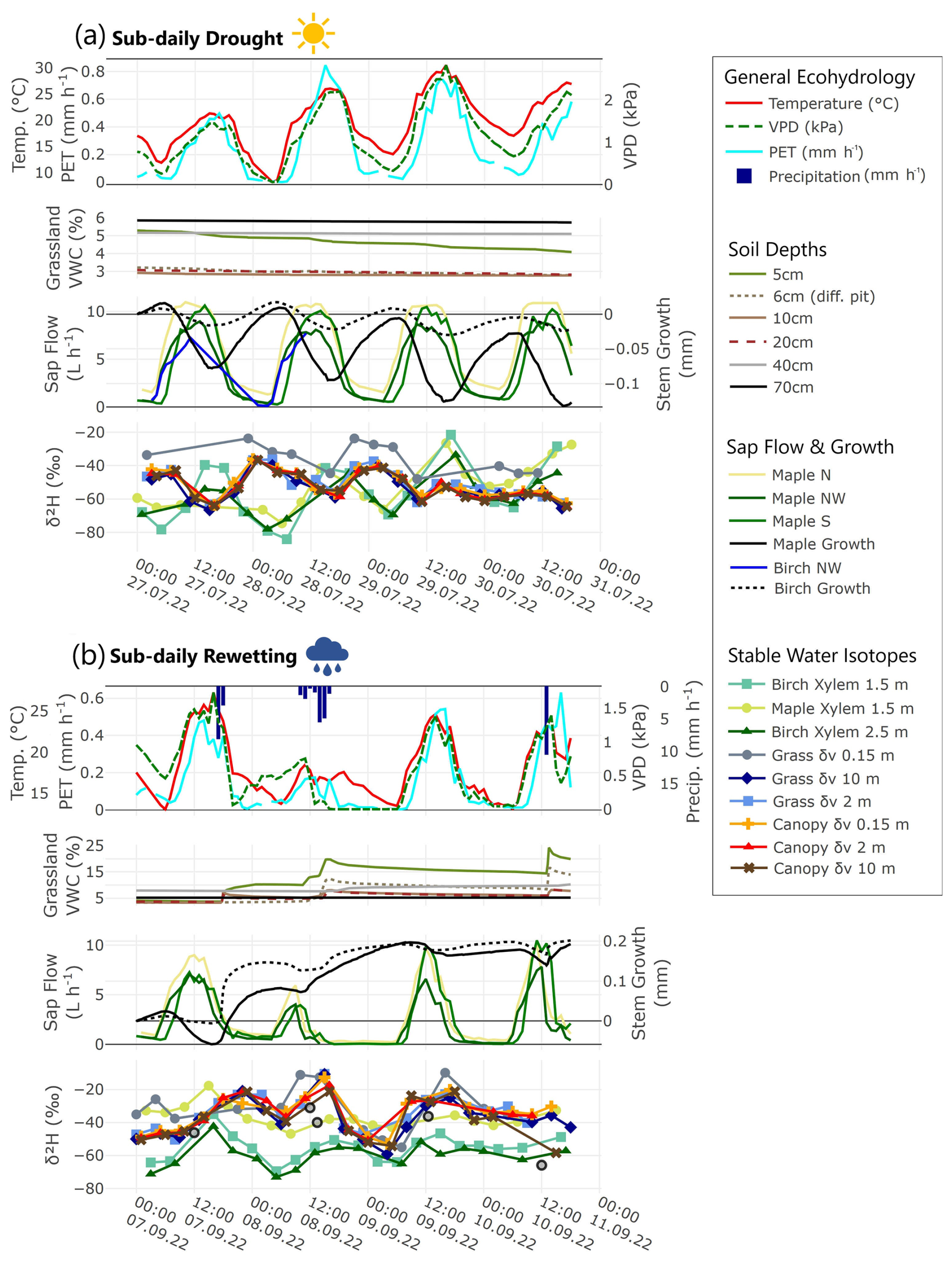
Figure 5Sub-daily timeseries displaying diurnal variation of tree xylem water (δxyl) and atmospheric vapor (δv) and measured ecohydrological variables during (a) drought conditions and (b) rewetting.
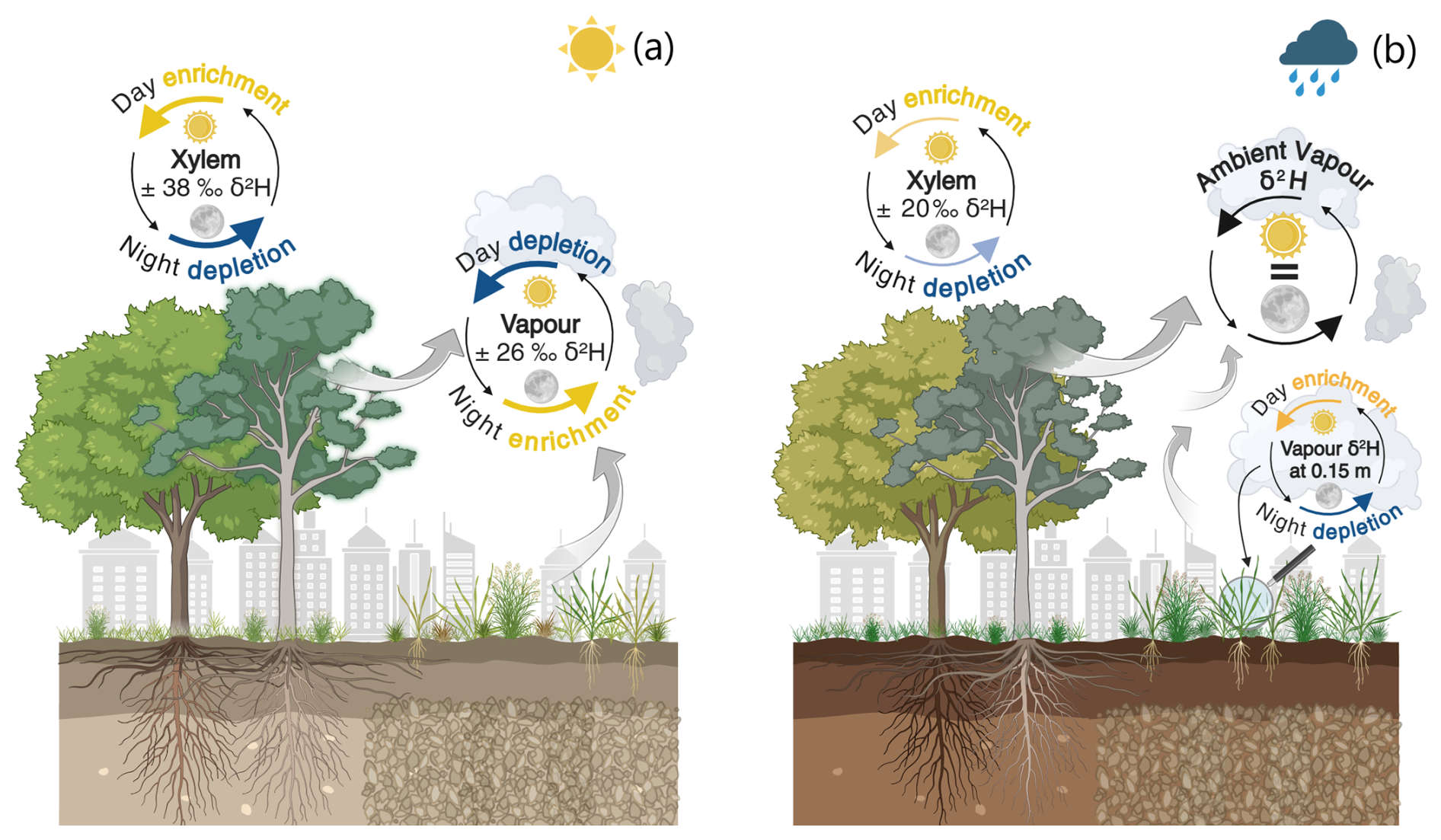
Figure 6Key figure summarizing the sub-daily change in signatures of stable water isotopes in urban tree xylem water (δxyl) and atmospheric water vapor (δv) (over grassland and under canopy). Day–night change defined as: day (08:00 a.m.–08:00 p.m. CEST) and night (08:00 p.m.–08:00 a.m. CEST). We compare summer drought (a) and autumn rewetting (b) in Berlin, Germany, 2022. Displayed amplitudes of the stable hydrogen isotope deuterium (δ2H [‰]) were calculated from hourly medians from the 24 h cycle of each period averaged over all δxyl and δv, respectively. Created in BioRender (CC BY-NC-ND), Ring (2025) https://BioRender.com/s1f2bm9 (last access: 12 August 2025).
Diurnal cycling of SWI became less distinct during the rewetting period (15 August–30 September 2022) (Fig. 4). Wilcoxon-tests no longer indicated significant sub-daily differences of δv values (p>0.05) during rewetting. Only δv values of grassland at 0.15 m (measured within higher grass) showed significant daytime enrichment (p=0.031). The sub-daily range of δxyl values averaged between all trees was smaller during rewetting (∼20 ‰ δ2H; Fig. 6), but an afternoon enrichment was still evident and differences statistically significant (p<0.05) revealed by Wilcoxon-tests.
In Fig. 5a we zoom into a typical 4 d long sub-daily timeseries (27 July–31 July 2022) of the monitored SWI and ecohydrological data during the drought period. Meteorological conditions 30 d prior to 27 July 2022 (Fig. S4a) were characterized by a daily mean temperature of 20.5 °C, a mean PET of 0.19 mm h−1 and accumulated precipitation of 33.4 mm (with 18.6 mm from a single storm event on 7 July). During the 4 d, sub-daily dynamics of temperature, VPD and PET were clearly radiation driven (daily maxima between 500–900 W m−2) with no precipitation. Soil moisture remained relatively constant over the 4 d period; only the upper soil layer (5 cm) became drier. Sap flow rates of both trees rose from morning hours until noon (up to 12 L h−1) and then decreased, dropping below 2.5 L h−1 during nighttime. Stem growth of both trees showed a similar temporal trend in a circadian cycle of daytime shrinking and nighttime swelling. The timeseries during drought showed clear opposite sub-daily cycling of δxyl and δv in δ2H values similar to the averages presented in Fig. 3. δxyl values became enriched in heavy isotopes during afternoons and depleted during nighttime (large sub-daily variation of ∼40 ‰ δ2H). The amplitudes in δxyl were greater in the lower borehole of B. pendula at 1.5 m compared to 2.5 m, which might be a dilution effect with height. δv values became enriched during nighttime hours (sub-daily variation of ∼20 ‰ δ2H). Lc-excess values are displayed in Fig. S4. Lc-excess of δv values was mostly above zero (∼5 ‰), with a single change to negative values during night on 28 July. δxyl value lc-excess was strongly negative with B. pendula lc-excess becoming more negative (around −20 ‰) than A. platanoides lc-excess (around −8 °). The drop in the lc-excess values after 28.7 at 00:00 can be explained by stronger entrainment processes during that night, meaning an intensified turbulent flux of water vapor that occurred between the relatively dry air in the free troposphere above and the moister air within the surface boundary layer (cf. Lai and Ehleringer, 2011; Lee et al., 2006).
In Fig. 5b we additionally zoom into a typical 4 d long sub-daily timeseries (7–11 September 2022) during the rewetting period. Meteorological conditions 30 d prior 7 September 2022 (Fig. S4b) were characterized by a daily mean temperature of 21 °C, a mean PET of 0.1 mm h−1 and a precipitation sum of 51.6 mm. During the 4 d, daily maxima of temperature, VPD and PET were dampened during the 7 h precipitation event on 8 September, but not during the shorter convectional events. Soil moisture rates indicated infiltration of precipitation to a depth of 5–20 cm in the sandy soil, especially after the longer 7 h rainfall. Soil moisture in depths below 30 cm was unaffected. Sap flow rates of the A. platanoides rose up to 10 L h−1 by noon. But also here, rates were dampened on the colder, rainy day of 8 September. Stem increments of both trees were picking up the precipitation signals consistent with a swelling of the bark.
The timeseries after two medium precipitation events showed smaller sub-daily variation of δxyl values (∼20 ‰ δ2H) compared to the presented drought timeseries which had no precipitation input. The two boreholes of birch showed similar amplitudes of δxyl values, implying no axial dampening during these days. δv signatures showed a drop in δ2H values after the rainfall events and stable values form the next morning onwards as indicated by the average rewetting period values from Fig. 4. Lc-excess values of δxyl and δv became higher compared to drought
The sub-daily timeseries during wet conditions (= after rewetting; 1–5 October 2022) (Fig. S5) showed lower maximum sap flow rates of A. platanoides. Soils were (re)wetted: Upper soils retained water from precipitation inputs for several days after a rain event. Lc-excess values of δv and δxyl during wet conditions showed overall positive values indicating lower influence of evaporative fractionation. Sub-daily variation of δxyl values was lowered compared to drought and rewetting (∼10 ‰ δ2H). δv signatures were influenced by the precipitation event.
Dual isotopes of all measured waters (Fig. S6, summary statistics in Tables S2 and S3, Supplement) revealed that deeper soil water and groundwater had isotopic signatures close to those of δv and δxyl. In contrast, dual isotopes of upper soil water and precipitation were more distinct from δv and δxyl dual isotope values. However, δv values of grassland at 0.15 m had similar signatures to upper soil waters. A comparison of the dual isotopes between the drought and rewetting period showed that δv (mean −44.79 ‰ δ2H and −7.45 ‰ δ18O) and δxyl (mean −55.02 ‰ δ2H and −7.09 ‰ δ18O) values corresponded more closely to the precipitation signatures (mean −43.37 ‰ δ2H and −6.72 ‰ δ18O)during the rewetting phase. Notably, δxyl values of B. pendula did not reflect the precipitation signal during rewetting. As the location of B. pendula was closer to a building and growing on a slope, we assume precipitation did not percolate as quick and soil replenishment was potentially composed of more fractionated waters.
4.3 Cross-Correlation Analysis of Sub-daily Isotopes and Ecohydrological Variables
A cross-correlation analysis between the monitored ecohydrological variables and in situ SWIvalues revealed some inversions of the sub-daily patterns between the drought and the rewetting period (Figs. S7, S8). Both δ2H and δ18O showed analogous patterns. During the summer drought, a high correlation of radiation () and an intermediate correlation of VPD () with δxyl and δv values was detected. It followed the diurnal radiation cycle with the inverse behavior of δxyl and δv signatures; δxyl had a positive correlation with solar radiation and VPD at lag 0 h. In contrast, δv values showed a negative correlation with solar radiation and VPD at lag 0 h. VPD maxima were lagged 2 h behind global radiation for both δxyl and δv values. This indicates that VPD had a delayed temporal influence on the isotope signatures than global radiation.
During rewetting, correlation signals of solar radiation and VPD with δxyl and δv values were damped: correlation coefficients were usually below (−)0.3. Lags of δv values above the grassland at 0.15 cm were intermediate between other heights of ambient vapor and tree xylem values during rewetting. Soil moisture at 30 cm depth below canopy showed only a low positive correlation (<0.3) with δxyl and δv values during drought. During rewetting, the correlation of soil moisture below the canopy with δxyl values stayed low but rose above 0.4 after a lag 0 h for δv values.
The cross-correlation between δxyl values of the A. platanoides borehole at 1.5 m and sap flow data of the A. platanoides (Figs. S9, S10) showed a positive correlation of 0.5 at a delay of ∼6 h during drought and a general radiation-driven sinusoidal curve. Stem growth and δxyl values of the A. platanoides (Figs. S9, S10) were only weakly correlated () during drought and showed the radiation driven sinusoidal curve. During rewetting, the correlation between δxyl values of A. platanoides and its sap flow was weaker. In addition, the correlation of δxyl values with stem growth stayed low but became positive indicating less radiation influence.
Cross-correlation analysis between δxyl values of the two B. pendula boreholes (1.5 and 2.5 m height; Figs. S9, S10) showed a high positive correlation (>0.5) at lag 0 h during both drought and rewetting with a sinusoidal radiation driven pattern. During drought, the maximum lag of the cross correlation between both boreholes of B. pendula had a delay of −1 h, which could hint at a travel time between both boreholes. During rewetting, there was a general shift to an overall positive correlation between the B. pendula boreholes and its stem growth. There was no significant correlation of B. pendula δxyl values with stem growth during drought, but an intermediate positive correlation of 0.33 at delay of −10 h during rewetting.
5.1 Diurnal water cycling in urban green spaces over the growing season
The combination of ecohydrological measurements and in situ monitoring of δv and δxyl signatures provided valuable long-term insights on diurnal water cycling processes in different urban vegetation over the growing season including summer drought conditions in 2022. We found significant sub-daily differences displayed in key Fig. 6 for both δv (amplitude ∼26 ‰ δ2H with daytime depletion) and δxyl (amplitude ∼38 ‰ δ2H with daytime enrichment) values during the drought. But during rewetting δv values showed constant signatures over the day. Uniquely δv values just above grassland (0.15 m) exhibited sub-daily cycling (amplitude ∼36 ‰ δ2H with significant daytime enrichment). Daytime enrichment of δxyl values was still significant during rewetting, but with reduced sub-daily amplitudes of ∼20 ‰ δ2H.
5.1.1 Ecohydrological fluxes
Through the integrated ecohydrological monitoring of urban trees (sap flow, LAI, stem growth, twig water potential), soil moisture and climate data we could identify drivers of distinct sub-daily variations in isotopic signatures of δv and δxyl of different green spaces to summer drought and autumn rewetting (Stevenson et al., 2023; Marx et al., 2022; Ring et al., 2024). During the drought period (1 July–14 August 2022), water cycling was predominantly radiation driven showing clear sub-daily variation (Fig. 5a). While sap flow rates peaked at noon, VPD reached the highest values in the afternoon for A. platanoides, hinting at stomatal control with increasing VPD or a radiation-VPD lag (O'Brien et al., 2004; Zhang et al., 2014). Peak sap flow rates were around 10 L h−1 suggesting no drought stress due to water supply by deeper water sources (cf. Ring et al., 2024). Mid-day values of twig water potential at the lower shaded canopy (Fig. S1) was most negative at −1.35 mPa, indicating no drop below the critical species-specific water potential value “P50” at which the plant experiences a 50 % loss of hydraulic conductivity at least for the lower canopy (Petruzzellis et al., 2022). A natural swelling of the tree stems was observed between midnight and morning hours, when VPD was low, followed by a shrinking during daytime as tree stems shrink and swell in relation to their water status (Steppe et al., 2015). Despite the precipitation input of 18.6 mm from a convectional event on 7 July – the water did not infiltrate into deeper soils (>30 cm depths), only replenishing upper soils for about a week until it was lost to ET fluxes. This persistence of dry soils further reduced ET processes enhancing moisture stress (cf. Kleine et al., 2020).
During the autumn rewetting period (15 August–30 September 2022), radiation inputs decreased and precipitation became the dominant factor influencing the water cycling within the SPAC. Higher twig water potential and swelling of the tree stems during rewetting revealed improved water supply (Herrmann et al., 2016; Schweiger et al., 2023). At the same time, PET rates (Fig. 2) decreased due to lowered radiation in autumn. Especially during precipitation events, temperature, VPD and PET rates were reduced (Fig. 5b). In parallel, we observed decreasing LAI and sap flow rates.
5.1.2 Atmospheric vapor isotopes below urban canopy and above urban grassland
The sub-daily isotopic signatures of in situ monitored δv values suggest systematic behaviour of diurnal water cycling in vegetated areas during summer drought. We detected clear daytime (08:00 a.m.–08:00 p.m. CEST) depletion in heavy isotopes of δv and vice versa and a night time enrichment at all heights under canopy and above grassland (Figs. 3, 6). As δv values were not correlated in time with soil moisture and also showed low coefficients in the cross-correlation with radiation and VPD (Figs. S7, S8) – we argue that during drought, not only ET processes were controlling δv values at daytime, but also atmospheric circulations of the larger urban surface boundary layer. A similar trend of diurnal cycling δv values during summer was found by in situ eddy covariance measurements of δv in the surface boundary layer of a forest in Germany (Braden-Behrens et al., 2019, 2020). They identified entrainment effects that were stronger than local ET as the main aspect for the daytime enrichment of δv in heavy isotopes. Also, Zannoni et al. (2025), identified that at sub-daily scales, vertical mixing is the dominant process affecting isotopic variability of δv values, which was high during drought in our study, when radiation was main influencing factor (Figs. S7, S8). This effect is related to mixing with dryer isotopically lighter δv values in the atmosphere surrounding the vegetation during daytime. This daytime entrainment results from the rapid molecular exchange of water isotopes between the moist surfaces and atmospheric water vapor, which can outweigh the impact of evaporation (Craig et al., 1963). At night time, the effect is reduced and local ET processes dominate, driving the isotopic enrichment. However, we argue that evaporation cannot be ruled out as a significant moisture source in the afternoon when the local convective boundary layer is stabilized (Lai and Ehleringer, 2011). At night, δv values partially exhibited negative lc-excess during the drought (Fig. S4a), emphasizing that the isotopic nocturnal enrichment of δv values is due to evaporative signals.
During the rewetting, we found no significant sub-daily differences (similar to Braden-Behrens et al., 2020) and an intermediate positive correlation with soil moisture for most of the measured heights of δv values (Figs. 4, S8). This indicates that δv values become more evaporation driven from the re-saturated upper soils (<30 cm depth), while entrainment is reduced with lower radiation (cf. Lai and Ehleringer, 2011). Only δv values monitored at a low height above the grass (0.15 m) showed significant daytime enrichment during rewetting (amplitude ∼36 ‰ δ2H with daytime enrichment). This suggests that ET fluxes were stronger just above the grass, likely promoted by higher moisture in the (enriched) upper soil water and at vegetation surfaces (cf. Ring et al., 2023). But it needs to be considered that for this valve, only 6.5 measurements were available on average to calculate the hourly median for each hour of the day during the rewetting period: we had to discard several measurements, which were probably due to increasing problems with condensation of cooler air in situ monitoring during autumn (cf. Landgraf et al., 2022). The overall positive lc-excess in δv values during rewetting (Fig. 5b) implies weaker evaporation signals occurred here, indicating that the re-wetted soils and the increasing number of precipitation events did not lead to higher E during autumn because of reduced radiation inputs. Angert et al. (2022) found that seasonal variations in near-surface δv values are driven by rainout effects, temperature and relative humidity controlling the initial vapor and variations in vertical mixing.
Thus, we assume that diurnal cycling of δv values over the growing season is driven by climatic and entrainment effects, while local ET does not fully dominate δv values during daytime, but also atmospheric factors within the footprint of the larger urban surface boundary layer. We identified an influence of local ET on δv values only at daytime just above grassland (0.15 m) during rewetting, when surfaces were very moist. Simple conceptual tracer-aided models, which integrated the high-resolution δv data from our study site, previously revealed that small-scale variations in interception and soil evaporation sources can subtly influence the composition of δv values (Birkel et al., 2025b). Considering that, sub-daily signals of δv have great potential to reveal previously unexplained short-term variances in SWI in daily resolved datasets to constrain conceptual models investigating the urban SPAC (cf. Birkel et al., 2025b).
5.1.3 Tree xylem waters
The relatively small sample size of two trees reflects the trade-off between measurement depth and spatial replication inherent to high-resolution in situ monitoring of δxyl values. Despite this, the temporal dynamics this study captured offer valuable perspectives on tree internal water cycling. The sub-daily isotopic signatures of in situ monitored δxyl values suggest systematic behaviour of sub-daily water cycling in urban trees. During summer drought, we detected strong differences between day- and night-time (08:00 p.m.–08:00 a.m. CEST) signatures of δxyl (amplitude ∼38 ‰ δ2H; Figs. 3, 6). In contrast to δv values, we identified an afternoon enrichment in heavy isotopes of δxyl when VPD and sap flow rates reached their maxima. The cross-correlation analysis revealed a dominant temporal connection between the diurnal hydroclimatic cycles of radiation and VPD with δxyl values during drought (Figs. S7, S8). While sap flow rates were highest at noon, maximum δxyl enrichment in heavy isotopes was ∼ 03:00 p.m. CEST (Fig. 3), like VPD maxima. Cross-correlation analysis also revealed a 1 h lag between the two boreholes of B. pendula during drought (Figs. S9, S10). As sap velocities were quite high, this seems less likely to indicate a 1 h travel time between boreholes, but rather an artefact of the sequential in situ measurements. But the amplitudes in δxyl day-time enrichment were occasionally larger in the lower borehole of B. pendula at 1.5 m compared to 2.5 m height, which might be a dilution effect with height. A recent in situ study by Minick et al. (2025) on internal water movement in oak tree xylem, saw slow axial advection and a loss of applied tracers in the shallow sap wood and with stem height, indicative of dilution and dampening with height. Likewise, De Deurwaerder et al. (2020) revealed such dynamic temporal fluctuations in measured δxyl values dependent on sampling height, which needs to be considered in sampling protocols.
While radiation inputs and PET were greatly reduced during rewetting, we still found significant day-time enrichment and night-time depletion in heavy isotopes of δxyl consistent with diurnal hydroclimatic cycling (Figs. 4, S7). But the sub-daily δxyl amplitudes of ∼20 ‰ δ2H were almost half smaller compared to the drought period (Fig. 6). Lc-excess of δxyl values became positive during the rewetting period, indicating a lowered evaporation signal in the xylem. Kühnhammer et al. (2022) likewise observed a clear daytime enrichment in heavy isotopes of δxyl in tropical rainforest trees with maxima around noon, and depleted δxyl values after sunset (sub-daily amplitude approx. 30 ‰ δ2H).
It exist various potential plant physiological processes driving daytime enrichment of δxyl values: During daytime, water demand in the tree canopy exceeds water absorption through the roots (Herzog et al., 1995; Cermák et al., 2007), which was reflectedin our study by the more negative twig water potential at noon compared to morning (Fig. S1). King et al. (2013) also reported a daily cycling of water uptake and loss in stem diameters of conifers in Switzerland, with the smallest diameters, occurring in the late evening and a diurnal amplitude that increased with temperature. In a 24 h field experiment including manual sampling of Eucalyptus globulus leaves, Cernusak et al. (2005) found the strongest enrichment of leaf water in the afternoon, while photosynthetic rates were highest in morning indicating that photosynthesis is not a dominant driver of diurnal tree SWI values. Most importantly, De Deurwaerder et al. (2020) list that the regulation of plant transpiration by stomata and root water uptake, which involves typical diurnal patterns linked to VPD, can have a sub-daily impact on δxyl values. In a lab experiment, Martín-Gómez et al. (2017) found short-term dynamics of δxyl signatures in response to drought, reflected in a xylem evaporative enrichment under limited sap flow. This is similar to our finding that maximum enrichment of δxyl values (∼ 03:00 p.m. CEST) does not follow maximum sap flow (noon) in time. Also, water loss and evaporation through the tree bark could be a driver for day-time δxyl enrichment, but is dependent on tree species and reduced with bark thickness (Dawson and Ehleringer, 1993; Lintunen et al., 2021; Zhou et al., 2024). Kübert et al. (2022) monitored δ-values of tree transpiration (δT) in situ and found that sub-daily cycling of δT-values was closely related to the diurnal variation of stomatal conductance. However, they discovered differences between in situ derived δT values and cryogenically extracted δxyl values, which hints for a possible deviation between δT and δxyl values. Earlier investigations indicate stomatal conductance as a driver of δT values but simultaneous non-steady-state effects within SWI fluxes of plants and isotopic fractionation leading to a variation between δT and δxyl values (Dubbert et al., 2017; Simonin et al., 2013). Additionally, it was found that δ2H and δ18O values in leaf water respond differently to environmental drivers (Cernusak et al., 2022). Finally, we argue that trees actively react and adapt to environmental conditions to moderate their water use throughout the day, which in turn influences δxyl values. Still, δxyl values likely differ from δT values due to complex physiological and environmental interactions. Key influencing factors include changes in radiation and VPD, as well as resulting shifts in sap flow dynamics, plant water potential, stomatal conductance and bark transpiration.
Our observations were made for deciduous species in temperate regions, but there is evidence that sub-daily cycling of δxyl is alternating within tree species and climate. A 24 h monitoring of rainforest tree species suggested that diurnal variation of δxyl is species-specific – some trees showed clear sub-daily differences, some not (Sohel et al., 2023b). This could be explained by sub-daily variations in leaf hydraulic conductance within species (Lo Gullo et al., 2005). Zhao et al. (2016) found overall stable diurnal δxyl values, but an afternoon depletion in twig water as well as a general fractionation between stem and twig δxyl waters in Populus euphratica. Bernhard et al. (2024) found no significant sub-daily fluctuations of manually sampled xylem water and its δxyl values in spruce, oak and beech of a temperate forest, but variations for different stem heights. Still, a daytime enrichment in heavy isotopes of δxyl is plausible for various tree species, as evaporative influence resulting from higher gas exchange rates and high gradients of water potential from roots to canopy have an impact on δxyl values.
Before implementing δxyl data into modelling, the validation and quantification of vegetative processes influencing variations in δxyl values is essential (De Deurwaerder et al., 2020). Biases in δxyl data from isotopic fractionation as a natural process are ambiguous for use of such data in models and can exacerbate isotopic discrimination of root water uptake depths (Birkel et al., 2025b).
5.2 Effects of summer drought on urban green spaces
5.2.1 Atmospheric moisture cycling during drought
Periods of drought are expected to not only become more frequent but also have greater longevity (Petrova et al., 2024), potentially impacting urban vegetation. Wei et al. (2015) found that the diurnal amplitude of δv values is larger during periods of lower transpiration, emphasizing that significant differences of δv values between daytime and night time occur during periods of water stress and limited transpiration. These diurnal cycling patterns of δv values during drought can be further impacted by the effects of climate change, when humidity differences between the surface boundary layer and atmosphere above are magnified by dry-air entrainment (van Heerwaarden et al., 2009). Although isolated high-intensity rainfall events can occur during summer droughts and their frequency and intensity are projected to increase (Sobaga et al., 2024), they often bring limited soil moisture replenishment. When rainfall intensity exceeds the soil infiltration capacity, water is rapidly lost through surface runoff to nearby water bodies or ponded water may increase evaporation. This effect can also be pronounced in urban areas where soils can be compacted by human footfall and sandy soils may have low water storage capacity and drain rapidly (Drastig et al., 2011; Ferreira et al., 2021). Further, drought periods can alter water transport in grasslands by restricted soil water mixing and reduced transpiration (Radolinski et al., 2025).
5.2.2 Tree water cycling during drought
The insights from in situ monitoring of δxyl values revealed significant impact of summer drought, indicated by large amplitudes δxyl values in the sub-daily water cycling and linked responses in plant physiological processes (see Sect. 5.1 and 5.1.3). During drought conditions, both A. platanoides and B. pendula exhibited no significant positive correlation of δxyl values with stem growth (as a proxy for sap flow) over the course of the day (Figs. S9, S10) indicating water limitation also at night.
Previous isotopic studies revealed that during drought tree canopies exhibit signs of stress, affecting their overall health and cooling capacity (Kuhlemann et al., 2021). Stevenson et al. (2025) further highlighted a strong sensitivity of urban green space hydrological functioning to seasonal hydroclimatic changes and high suppression of ET fluxes in urban trees in SW-Berlin during the 2022 drought. Gessler et al. (2022) showed that drought reduces water uptake in beech from the drying topsoil, but also with no compensatory uptake from deeper soil layers. Such limitation can especially impair tree functioning when temperatures are high during daytime. Kupper et al. (2017) found that elevated daytime atmospheric humidity increases the potential for night time water flux in B. pendula. This effect is reduced under drought conditions, so that night-time recharge may also have been more constrained in the B. pendula analyzed.
Drought severity and antecedent climatic conditions have been shown to be major factors influencing the magnitude of sub-daily drought stress on vegetation. Kübert et al. (2022) found similar sub-daily cycling with daytime enrichment in δT values in trees of an enclosed rainforest, but here the amplitude of the diurnal variation strongly declined with increasing drought severity. In our study, the drought conditions during summer were less pronounced compared to their study: as the preceding 2021/22 winter was wet and some precipitation inputs were recorded during summer (Fig. 2). Possibly with more severe drought conditions less distinct diurnal cycling would be evident. Further, the effects of droughts on tree functioning depend on species composition and their adaptation to drought. It was previously reported that trees can be in competition for water with neighbouring species (Kinzinger et al., 2024; Magh et al., 2020).
To conclude, long summer droughts affect the sub-daily water cycling in tree xylem, which results in limited ET fluxes and reduced sap flow rates relative to high VPD rates during day time as well as a stagnation of night-time stem swelling. The magnitude of these effects is likely to be highly variable and dependent on the ecohydrological status of tree species composition within green spaces.
5.3 Wider implications
This study provided the opportunity to better understand the sub-daily dynamics of water cycling in different types of vegetation over the growing season. The insights of our high-resolution SWI dataset on plant physiology and hydrological functioning, including atmospheric responses are of high relevance for studying ecohydrological partitioning (Birkel et al., 2025b). SWI of plant xylem and the surrounding atmosphere revealed clear diurnal cycling patterns, reflecting changes in radiation and VPD. State of the art SWI isotope assessment should transition towards in situ sampling to better catch complex sub-daily root water uptake and plant internal water cycling patterns. Moreover, investigating species-specific responses of trees across diverse biomes is essential (Sohel et al., 2023b). As we only measured δxyl values, we suggest to include measurements of SWI covering the whole tree from root to stem and branches to twigs, including direct measurements of transpiration for future in situ studies on sub-daily tree water cycling (Sohel et al., 2023a). Nevertheless, in situ monitoring providing sub-daily datasets is not always possible due to difficult sampling locations, high maintenance requirements or cost of instruments. We suggest more appropriate δxyl sampling protocols (Tierney et al., 2025), that include sampling at the same time of day, preferably in morning hours after sunrise which mostly reflect a daily mean value. Mid-day and afternoon sampling of plant water might very likely reflect daytime enrichment and further fractionation in SWI. Finally, we join others in calling for an exhaustive evaluation of in situ methods vs extracted waters to close the questioning about the validity of in situ methods towards a common methodological framework of studies of the SPAC (cf. Ceperley et al., 2024).
Our high-resolution dataset also convenes a more quantitative understanding of ecohydrological fluxes and partitioning within green spaces, since the continuous, sub-daily (3-hourly) dataset of δv and δxyl values is highly valuable to constrain ecohydrological models which help predicting impacts of climate change on vegetation mediated water fluxes. The application of continuous water isotope data reduces parameter uncertainties in SPAC modelling (Smith et al., 2021). Birkel et al. (2025a) have just further emphasised the importance of including SWI in modelling to advance climate change prediction. Additionally, our ecohydrological dataset has the potential to upgrade the assumptions of remote sensing-based modelling of sub-daily water fluxes within vegetation (cf. Vermunt et al., 2022).
This study was able to capture vegetation water cycling during a summer drought, at a time when increasing drought and heatwaves with complex land-atmosphere feedbacks is a problem all over the world (van Loon, 2015; Miralles et al., 2019). Drier soils contribute to the intensification of heatwaves through reduced evaporative cooling and increased sensible heat fluxes, but at the same time ET processes can amplify the effects of negative precipitation anomalies (Teuling et al., 2013; Dirmeyer et al., 2021). These effects are further modulated by the type of land-use and soil: In European grasslands heating is on short-term suppressed be ET cooling, but at long term soil moisture depletion is accelerated underneath grasslands, whereas forests mitigate the impact of extreme heat more at long-term even though they have higher interception and transpiration losses (Teuling et al., 2010; Kleine et al., 2020). Our findings support vegetation management that prioritises plant physiological functioning and water storage within the landscape (Luo et al., 2024). Vegetation worldwide responds to drying surface soils by shifting water uptake to deeper layers, thereby sustaining transpiration and ensuring hydraulic safety during seasonal droughts (Bachofen et al., 2024). The trees investigated in our study were mature, deep-rooted A. platanoides and B. pendula individuals that accessed deeper soil water and groundwater for sustenance, ensuring sufficient sap flow rates and maintaining overall tree functioning (cf. Ring et al., 2024). However, smaller, younger trees with restricted rooting space and high surface sealing in cities face additional challenges and are more vulnerable to drought stress (Kluge and Kirmaier, 2024; Anys and Weiler, 2025). Therefore, it is important to choose climate resilient tree species that can sustain their natural water supply under drought (Roloff et al., 2009).
During the growing season of 2022, we combined in situ monitoring via CRDS and ecohydrological monitoring in different urban vegetation in Berlin, Germany. SWI of atmospheric water vapor (δv) were monitored at different heights above a grassland and under the canopy of a mature A. platanoides tree. SWI of the xylem water within tree stems (δxyl) was monitored at different heights in an A. platanoides and B. pendula tree.
The summer period from July until mid-August was characterised by drought conditions, including occasional convective rainfall events, which did not infiltrate into deeper soils, and was followed by a rewetting period with frequent rainfalls starting in mid-August. We identified distinct sub-daily ecohydrological dynamics over drought and rewetting. Throughout the drought period, water cycling was predominantly radiation driven, with highest VPD rates in the afternoons and persisting dry soils. During rewetting, precipitation became the dominant driver of the water cycling within the SPAC, while at the same time PET rates decreased.
The analysis of sub-daily isotopes suggests systematic behaviour of both δv and δxyl values during summer drought. δv values were characterized by a daytime depletion in heavy isotopes, driven by entrainment effects and evaporation. We found daytime enrichment in δxyl signatures, with maximum enrichment in afternoons, which was driven by VPD , stomatal regulation of transpiration and possibly bark transpiration. For the rewetting, we revealed that the systematic diurnal cycling of δv values was discontinued for most heights. It became more evaporation driven from the re-saturated soils, while the influence of entrainment from the larger urban surface boundary layer was reduced. Only δv values just above the grassland surface (0.15 m) showed a significant daytime enrichment (amplitude ∼36 ‰ δ2H), hinting for ET fluxes from vegetation surfaces and superficially enriched soil water. While radiation inputs were greatly reduced during rewetting, we still found significant daytime enrichment and night-time depletion of δxyl values with sub-daily amplitudes almost halved compared to the drought period. Our results suggest that the monitored urban vegetation experienced high water demand during summer drought. Constrained ET fluxes were indicated by reduced precipitation and soil moisture. In the trees, this was reflected in lowered sap flux relative to VPD in the afternoons, as well as stagnated night-time trunk swelling, but the mature trees could overall sustain their physiological functioning. Our findings provide novel insights on plant physiology and hydrological functioning by demonstrating the relevance of high-resolution SWI data and emphasizing the need for in situ SWI assessment to capture sub-daily plant water uptake and internal water cycling. Such continuous, sub-daily datasets of δv and δxyl values have the potential to better constrain ecohydrological models which help predicting impacts of climate change on plant mediated water fluxes. However, more research is needed to extend our findings on sub-daily water cycling within vegetated areas, covering different types of planting, higher sample size and varying tree species from root to twig, including leaf transpiration.
The dataset used in this study is subject to a three-year embargo following the expiry of DFG Project funding for Ann-Marie Ring and will therefore be made openly available on 1 December 2028 in the open-access database FRED at IGB. Until then, the isotope and ecohydrology data are available with a password (to be received upon request from Doerthe Tetzlaff) from the open-access database FRED at IGB – Data package No. 979.
The supplement related to this article is available online at https://doi.org/10.5194/hess-29-6663-2025-supplement.
AMR: Conceptualization, Data curation, Data analysis and plotting, Methodology, Field activities, Writing – original draft preparation. DT: Investigation, Funding acquisition, Data curation, Methodology, Formal analysis, Writing – review & editing. CB: Investigation, Methodology, Formal analysis, Writing – review & editing. CS: Conceptualization, Investigation, Methodology, Formal analysis, Writing – review & editing.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank all colleagues involved in installing the in situ monitoring setup and sampling precipitation, groundwater, and soil water isotopes, in particular David Dubbert, Jessica Landgraf, Jonas Freymüller, Hauke Dämpfling, Jan Christopher, and Lisa Grof, as well as Grit Siegert (analysis of soil particle size) and Johanna Stralendorff (twig water potential and LAI measurements).
This research has been supported by the German Research Foundation (DFG) as part of the Research Training Group “Urban Water Interfaces” (UWI; GRK2032/2) and the Einstein Foundation as part of the “Modelling surface and groundwater with isotopes in urban catchments” (MOSAIC) project. Funding for DT was also received through the Einstein Research Unit “Climate and Water under Change” from the Einstein Foundation Berlin and the Berlin University Alliance (grant no. ERU-2020-609), the BiNatur project (BMBF no. 16LW0156), and the “Wasserressourcenpreis 2024” of the Rüdiger Kurt Bode-Foundation. We also acknowledge the BMBF (funding code 033W034A), which supported the stable isotope laboratory and in situ laser analyser. Further, Christian Birkel’s contribution was funded by a senior fellowship of the Leibniz Institute of Freshwater Ecology and Inland Fisheries (IGB) and a sabbatical license from UCR. Chris Soulsby was additionally funded by the Leverhulme Trust through the ISOLAND project (grant no. RPG 2018 375).
This open-access publication was funded by the Humboldt-Universität zu Berlin.
This paper was edited by Miriam Coenders-Gerrits and reviewed by two anonymous referees.
Allen, R. G., Pereira, L. S., Raes, D., and Smith, M.: Crop evapotranspiration - Guidelines for computing crop water requirements, FAO Irrigation and Drainage, 56, ISBN 92-5-104219-5, 1998.
Angert, A., Lee, J.-E., and Yakir, D.: Seasonal variations in the isotopic composition of near-surface water vapour in the eastern Mediterranean, Tellus B, 60, 674, https://doi.org/10.1111/j.1600-0889.2008.00357.x, 2022.
Anys, M. and Weiler, M.: Drought Impact on Transpiration Dynamics of Common Deciduous Trees Growing at Contrasting Urban Sites, Ecohydrology, 18, https://doi.org/10.1002/eco.70007, 2025.
Asbjornsen, H., Goldsmith, G. R., Alvarado-Barrientos, M. S., Rebel, K., van Osch, F. P., Rietkerk, M., Chen, J., Gotsch, S., Tobon, C., Geissert, D. R., Gomez-Tagle, A., Vache, K., and Dawson, T. E.: Ecohydrological advances and applications in plant-water relations research: a review, Journal of Plant Ecology, 4, 3–22, https://doi.org/10.1093/jpe/rtr005, 2011.
Bachofen, C., Tumber-Dávila, S. J., Mackay, D. S., McDowell, N. G., Carminati, A., Klein, T., Stocker, B. D., Mencuccini, M., and Grossiord, C.: Tree water uptake patterns across the globe, The New phytologist, 242, 1891–1910, https://doi.org/10.1111/nph.19762, 2024.
Barbeta, A., Gimeno, T. E., Clavé, L., Fréjaville, B., Jones, S. P., Delvigne, C., Wingate, L., and Ogée, J.: An explanation for the isotopic offset between soil and stem water in a temperate tree species, The New phytologist, 227, 766–779, https://doi.org/10.1111/nph.16564, 2020.
Beck, H. E., Zimmermann, N. E., McVicar, T. R., Vergopolan, N., Berg, A., and Wood, E. F.: Present and future Köppen-Geiger climate classification maps at 1-km resolution, Scientific data, 5, 180214, https://doi.org/10.1038/sdata.2018.214, 2018.
Bernacchi, C. J. and VanLoocke, A.: Terrestrial ecosystems in a changing environment: a dominant role for water, Annual review of plant biology, 66, 599–622, https://doi.org/10.1146/annurev-arplant-043014-114834, 2015.
Bernhard, F., Floriancic, M. G., Treydte, K., Gessler, A., Kirchner, J. W., and Meusburger, K.: Tree- and stand-scale variability of xylem water stable isotope signatures in mature beech, oak and spruce, Ecohydrology, 17, https://doi.org/10.1002/eco.2614, 2024.
Beyer, M., Kühnhammer, K., and Dubbert, M.: In situ measurements of soil and plant water isotopes: a review of approaches, practical considerations and a vision for the future, Hydrol. Earth Syst. Sci., 24, 4413–4440, https://doi.org/10.5194/hess-24-4413-2020, 2020.
Birkel, C., Miller, J., Watson, A., Anh Trinh, D., Durán-Quesada, A. M., Sánchez-Murillo, R., Soulsby, C., Terzer-Wassmuth, S., Tetzlaff, D., Uhlenbrook, S., Vystavna, Y., and Yoshimura, K.: Demystifying the art of isotope-enabled hydrological and climate modelling, The Science of the total environment, 959, 178242, https://doi.org/10.1016/j.scitotenv.2024.178242, 2025a.
Birkel, C., Tetzlaff, D., Ring, A.-M., and Soulsby, C.: Does high resolution in situ xylem and atmospheric vapor isotope data help improve modeled estimates of ecohydrological partitioning?, Agricultural and Forest Meteorology, 365, 110467, https://doi.org/10.1016/j.agrformet.2025.110467, 2025b.
Bliss Singer, M., Asfaw, D. T., Rosolem, R., Cuthbert, M. O., Miralles, D. G., MacLeod, D., Quichimbo, E. A., and Michaelides, K.: Hourly potential evapotranspiration at 0.1° resolution for the global land surface from 1981-present, Scientific data, 8, 224, https://doi.org/10.1038/s41597-021-01003-9, 2021.
Bonferroni, C. E.: Il calcolo delle assicurazioni su gruppi di teste, Studi in Onore del Professore Salvatore Ortu Carboni, Rome, Italy, 13–60, 1935.
Braden-Behrens, J., Markwitz, C., and Knohl, A.: Eddy covariance measurements of the dual-isotope composition of evapotranspiration, Agricultural and Forest Meteorology, 269–270, 203–219, https://doi.org/10.1016/j.agrformet.2019.01.035, 2019.
Braden-Behrens, J., Siebicke, L., and Knohl, A.: Drivers of the variability of the isotopic composition of water vapor in the surface boundary layer, Biogeosciences Discuss [preprint], https://doi.org/10.5194/bg-2020-398, 2020.
Ceperley, N., Gimeno, T. E., Jacobs, S. R., Beyer, M., Dubbert, M., Fischer, B., Geris, J., Holko, L., Kübert, A., Le Gall, S., Lehmann, M. M., Llorens, P., Millar, C., Penna, D., Prieto, I., Radolinski, J., Scandellari, F., Stockinger, M., Stumpp, C., Tetzlaff, D., van Meerveld, I., Werner, C., Yildiz, O., Zuecco, G., Barbeta, A., Orlowski, N., and Rothfuss, Y.: Toward a common methodological framework for the sampling, extraction, and isotopic analysis of water in the Critical Zone to study vegetation water use, WIREs Water, 11, https://doi.org/10.1002/wat2.1727, 2024.
Cermák, J., Kucera, J., Bauerle, W. L., Phillips, N., and Hinckley, T. M.: Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas-fir trees, Tree physiology, 27, 181–198, https://doi.org/10.1093/treephys/27.2.181, 2007.
Cernusak, L. A., Farquhar, G. D., and Pate, J. S.: Environmental and physiological controls over oxygen and carbon isotope composition of Tasmanian blue gum, Eucalyptus globulus, Tree physiology, 25, 129–146, https://doi.org/10.1093/treephys/25.2.129, 2005.
Cernusak, L. A., Barbeta, A., Bush, R. T., Eichstaedt Bögelein, R., Ferrio, J. P., Flanagan, L. B., Gessler, A., Martín-Gómez, P., Hirl, R. T., Kahmen, A., Keitel, C., Lai, C.-T., Munksgaard, N. C., Nelson, D. B., Ogée, J., Roden, J. S., Schnyder, H., Voelker, S. L., Wang, L., Stuart-Williams, H., Wingate, L., Yu, W., Zhao, L., and Cuntz, M.: Do 2 H and 18 O in leaf water reflect environmental drivers differently?, The New phytologist, 235, 41–51, https://doi.org/10.1111/nph.18113, 2022.
Chambers, J., Eddy, W., Härdle, W., Sheather, S., Tierney, L., Venables, W. N., and Ripley, B. D.: Modern Applied Statistics with S, Springer New York, New York, NY, 2002.
Craig, H., Gordon, L. I., and Horibe, Y.: Isotopic exchange effects in the evaporation of water: 1. Low-temperature experimental results, J. Geophys. Res., 68, 5079–5087, https://doi.org/10.1029/JZ068i017p05079, 1963.
Dahlmann, A., Marshall, J. D., Dubbert, D., Hoffmann, M., and Dubbert, M.: Simple water vapor sampling for stable isotope analysis using affordable valves and bags, Atmos. Meas. Tech., 18, 2607–2618, https://doi.org/10.5194/amt-18-2607-2025, 2025.
Dawson, T. E. and Ehleringer, J. R.: Isotopic enrichment of water in the “woody” tissues of plants: Implications for plant water source, water uptake, and other studies which use the stable isotopic composition of cellulose, Geochimica et Cosmochimica Acta, 57, 3487–3492, https://doi.org/10.1016/0016-7037(93)90554-A, 1993.
De Deurwaerder, H. P. T., Visser, M. D., Detto, M., Boeckx, P., Meunier, F., Kuehnhammer, K., Magh, R.-K., Marshall, J. D., Wang, L., Zhao, L., and Verbeeck, H.: Causes and consequences of pronounced variation in the isotope composition of plant xylem water, Biogeosciences, 17, 4853–4870, https://doi.org/10.5194/bg-17-4853-2020, 2020.
Dirmeyer, P. A., Balsamo, G., Blyth, E. M., Morrison, R., and Cooper, H. M.: Land-Atmosphere Interactions Exacerbated the Drought and Heatwave Over Northern Europe During Summer 2018, AGU Advances, 2, https://doi.org/10.1029/2020AV000283, 2021.
Drastig, K., Prochnow, A., Baumecker, M., Berg, W., and Brunsch, R.: Agricultural Water Management in Brandenburg, DIE ERDE – Journal of the Geographical Society of Berlin, 142, 119–140, 2011.
Dubbert, M. and Werner, C.: Water fluxes mediated by vegetation: emerging isotopic insights at the soil and atmosphere interfaces, The New phytologist, 221, 1754–1763, https://doi.org/10.1111/nph.15547, 2019.
Dubbert, M., Kübert, A., and Werner, C.: Impact of Leaf Traits on Temporal Dynamics of Transpired Oxygen Isotope Signatures and Its Impact on Atmospheric Vapor, Frontiers in plant science, 8, 5, https://doi.org/10.3389/fpls.2017.00005, 2017.
DWD: GPCC Drought Index July 2022, https://www.dwd.de/DE/leistungen/rcccm/int/rcccm_int_spi.html (last access: 14 August 2025), 2022a.
DWD: Deutschlandwetter im Frühling 2022, https://www.dwd.de/DE/presse/pressemitteilungen/DE/2022/20220429_deutschlandwetter_fruehling2022_news.html (last access: 13 August 2025), 2022b.
DWD: Deutschlandwetter im Sommer 2022, https://www.dwd.de/DE/presse/pressemitteilungen/DE/2022/20220830_deutschlandwetter_sommer2022_news.html (last access: 13 August 2025), 2022c.
DWD: Multi-year averages for reference period 1991–2020, https://www.dwd.de/DE/leistungen/klimadatendeutschland/vielj_mittelwerte.html (last access: 13 August 2025), 2023.
DWD: Tägliche Stationsbeobachtungen (Temperatur, Druck, Niederschlag, Sonnenscheindauer, etc.) für Deutschland; Version v24.3, https://opendata.dwd.de/climate_environment/CDC/observations_germany/climate/daily/kl/recent/ (last access: 13 August 2025), 2024.
Ferreira, C. S., Kalantari, Z., Seifollahi-Aghmiuni, S., Ghajarnia, N., Rahmati, O., and Solomun, M. K.: Rainfall-runoff-erosion processes in urban areas, in: Precipitation, edited by: Rodrigo-Comino, J., Elsevier, 481–498, https://doi.org/10.1016/B978-0-12-822699-5.00018-5, 2021.
Friedman, M.: The Use of Ranks to Avoid the Assumption of Normality Implicit in the Analysis of Variance, Journal of the American Statistical Association, 32, 675, https://doi.org/10.2307/2279372, 1937.
Galewsky, J., Steen-Larsen, H. C., Field, R. D., Worden, J., Risi, C., and Schneider, M.: Stable isotopes in atmospheric water vapor and applications to the hydrologic cycle, Rev. Geophys., 54, 809–865, https://doi.org/10.1002/2015RG000512, 2016.
Geoportal Berlin: Soil types, https://fbinter.stadt-berlin.de/fb/index.jsp (last access: 15 January 2025), 2015.
Gessler, A., Bächli, L., Rouholahnejad Freund, E., Treydte, K., Schaub, M., Haeni, M., Weiler, M., Seeger, S., Marshall, J., Hug, C., Zweifel, R., Hagedorn, F., Rigling, A., Saurer, M., and Meusburger, K.: Drought reduces water uptake in beech from the drying topsoil, but no compensatory uptake occurs from deeper soil layers, The New phytologist, 233, 194–206, https://doi.org/10.1111/nph.17767, 2022.
Haberstroh, S., Kübert, A., and Werner, C.: Two common pitfalls in the analysis of water-stable isotopologues with cryogenic vacuum extraction and cavity ring-down spectroscopy, Analytical science advances, 5, 2300053, https://doi.org/10.1002/ansa.202300053, 2024.
Helliker, B. R., Roden, J. S., Cook, C., and Ehleringer, J. R.: A rapid and precise method for sampling and determining the oxygen isotope ratio of atmospheric water vapor, Rapid communications in mass spectrometry RCM, 16, 929–932, https://doi.org/10.1002/rcm.659, 2002.
Herbstritt, B., Gralher, B., Seeger, S., Rinderer, M., and Weiler, M.: Technical note: Discrete in situ vapor sampling for subsequent lab-based water stable isotope analysis, Hydrol. Earth Syst. Sci., 27, 3701–3718, https://doi.org/10.5194/hess-27-3701-2023, 2023.
Herrmann, V., McMahon, S. M., Detto, M., Lutz, J. A., Davies, S. J., Chang-Yang, C.-H., and Anderson-Teixeira, K. J.: Tree Circumference Dynamics in Four Forests Characterized Using Automated Dendrometer Bands, PloS one, 11, e0169020, https://doi.org/10.1371/journal.pone.0169020, 2016.
Herzog, K., Hsler, R., and Thum, R.: Diurnal changes in the radius of a subalpine Norway spruce stem: their relation to the sap flow and their use to estimate transpiration, Trees, 10, https://doi.org/10.1007/BF00192189, 1995.
Hughes, C. E. and Crawford, J.: A new precipitation weighted method for determining the meteoric water line for hydrological applications demonstrated using Australian and global GNIP data, Journal of Hydrology, 464–465, 344–351, https://doi.org/10.1016/j.jhydrol.2012.07.029, 2012.
Khaliq, M. N., Ouarda, T., Gachon, P., Sushama, L., and St-Hilaire, A.: Identification of hydrological trends in the presence of serial and cross correlations: A review of selected methods and their application to annual flow regimes of Canadian rivers, Journal of Hydrology, 368, 117–130, https://doi.org/10.1016/j.jhydrol.2009.01.035, 2009.
Kim, Y., Garcia, M., Morillas, L., Weber, U., Black, T. A., and Johnson, M. S.: Relative humidity gradients as a key constraint on terrestrial water and energy fluxes, Hydrol. Earth Syst. Sci., 25, 5175–5191, https://doi.org/10.5194/hess-25-5175-2021, 2021.
King, G., Fonti, P., Nievergelt, D., Büntgen, U., and Frank, D.: Climatic drivers of hourly to yearly tree radius variations along a 6°C natural warming gradient, Agricultural and Forest Meteorology, 168, 36–46, https://doi.org/10.1016/j.agrformet.2012.08.002, 2013.
Kinzinger, L., Mach, J., Haberstroh, S., Schindler, Z., Frey, J., Dubbert, M., Seeger, S., Seifert, T., Weiler, M., Orlowski, N., and Werner, C.: Interaction between beech and spruce trees in temperate forests affects water use, root water uptake pattern and canopy structure, Tree physiology, 44, https://doi.org/10.1093/treephys/tpad144, 2024.
Kleine, L., Tetzlaff, D., Smith, A., Wang, H., and Soulsby, C.: Using water stable isotopes to understand evaporation, moisture stress, and re-wetting in catchment forest and grassland soils of the summer drought of 2018, Hydrol. Earth Syst. Sci., 24, 3737–3752, https://doi.org/10.5194/hess-24-3737-2020, 2020.
Kluge, B. and Kirmaier, M.: Urban trees left high and dry – Modelling urban trees water supply and evapotranspiration under drought, Environ. Res. Commun., 6, 115029, https://doi.org/10.1088/2515-7620/ad7dda, 2024.
Konarska, J., Uddling, J., Holmer, B., Lutz, M., Lindberg, F., Pleijel, H., and Thorsson, S.: Transpiration of urban trees and its cooling effect in a high latitude city, International journal of biometeorology, 60, 159–172, https://doi.org/10.1007/s00484-015-1014-x, 2016.
Kübert, A., Dubbert, M., Bamberger, I., Kühnhammer, K., Beyer, M., van Haren, J., Bailey, K., Hu, J., Meredith, L. K., Nemiah Ladd, S., and Werner, C.: Tracing plant source water dynamics during drought by continuous transpiration measurements: An in-situ stable isotope approach, Plant, cell & environment, 46, 133–149, https://doi.org/10.1111/pce.14475, 2022.
Kuhlemann, L.-M., Tetzlaff, D., Smith, A., Kleinschmit, B., and Soulsby, C.: Using soil water isotopes to infer the influence of contrasting urban green space on ecohydrological partitioning, Hydrol. Earth Syst. Sci., 25, 927–943, https://doi.org/10.5194/hess-25-927-2021, 2021.
Kühnhammer, K., Dahlmann, A., Iraheta, A., Gerchow, M., Birkel, C., Marshall, J. D., and Beyer, M.: Continuous in situ measurements of water stable isotopes in soils, tree trunk and root xylem: Field approval, Rapid Communications in Mass Spectrometry, 36, e9232, https://doi.org/10.1002/rcm.9232, 2022.
Kupper, P., Rohula, G., Inno, L., Ostonen, I., Sellin, A., and Sõber, A.: Impact of high daytime air humidity on nutrient uptake and night-time water flux in silver birch, a boreal forest tree species, Reg. Environ. Change, 17, 2149–2157, https://doi.org/10.1007/s10113-016-1092-2, 2017.
Lai, C.-T. and Ehleringer, J. R.: Deuterium excess reveals diurnal sources of water vapor in forest air, Oecologia, 165, 213–223, https://doi.org/10.1007/s00442-010-1721-2, 2011.
Landgraf, J., Tetzlaff, D., Dubbert, M., Dubbert, D., Smith, A., and Soulsby, C.: Xylem water in riparian willow trees (Salix alba) reveals shallow sources of root water uptake by in situ monitoring of stable water isotopes, Hydrol. Earth Syst. Sci., 26, 2073–2092, https://doi.org/10.5194/hess-26-2073-2022, 2022.
Landwehr, J. M. and Coplen, T.: Line-conditioned excess: a new method for characterizing stable hydrogen and oxygen isotope ratios in hydrologic systems, in: Isotopes in environmental studies - Aquatic Forum 2004, International Atomic Energy Agency (IAEA), ISBN 92-0-111305-X, 2006.
Lee, X., Smith, R., and Williams, J.: Water vapour 18O/16O isotope ratio in surface air in New England, USA, Tellus B, 58, 293, https://doi.org/10.1111/j.1600-0889.2006.00191.x, 2006.
Limberg, A., Darkow, P., Faensen-Thiebes, A., Fritz-Taute, B., Günther, M., Hähnel, K., and Hörmann, U., Jahn, D., Köhler, A. Krüger, E., May, S., Naumann, J., and Wagner, M.: Grundwasser in Berlin, Vorkommen ⋅ Nutzung ⋅ Schutz ⋅ Gefährdung, Senatsverwaltung für Gesundheit, Umwelt und Verbraucherschutz, Berlin, https://www.berlin.de/sen/uvk/_assets/umwelt/wasser-und-geologie/publikationen-und-merkblaetter/grundwasser-broschuere.pdf (last access: 15 January 2025), 2007.
Lintunen, A., Preisler, Y., Oz, I., Yakir, D., Vesala, T., and Hölttä, T.: Bark Transpiration Rates Can Reach Needle Transpiration Rates Under Dry Conditions in a Semi-arid Forest, Frontiers in plant science, 12, 790684, https://doi.org/10.3389/fpls.2021.790684, 2021.
Lo Gullo, M. A., Nardini, A., Trifilò, P., and Salleo, S.: Diurnal and seasonal variations in leaf hydraulic conductance in evergreen and deciduous trees, Tree physiology, 25, 505–512, https://doi.org/10.1093/treephys/25.4.505, 2005.
Luo, S., Tetzlaff, D., Smith, A., and Soulsby, C.: Long-term drought effects on landscape water storage and recovery under contrasting landuses, Journal of Hydrology, 636, 131339, https://doi.org/10.1016/j.jhydrol.2024.131339, 2024.
Lüthgens, C. and Böse, M.: Chronology of Weichselian main ice marginal positions in north-eastern Germany, E&G Quaternary Sci. J., 60, 236–247, https://doi.org/10.3285/eg.60.2-3.02, 2011.
Magh, R.-K., Eiferle, C., Burzlaff, T., Dannenmann, M., Rennenberg, H., and Dubbert, M.: Competition for water rather than facilitation in mixed beech-fir forests after drying-wetting cycle, Journal of Hydrology, 587, 124944, https://doi.org/10.1016/j.jhydrol.2020.124944, 2020.
Majoube, M.: Fractionnement en oxygène 18 et en deutérium entre l'eau et sa vapeur, J. Chim. Phys., 68, 1423–1436, https://doi.org/10.1051/jcp/1971681423, 1971.
Mann, H. B. and Whitney, D. R.: On a Test of Whether one of Two Random Variables is Stochastically Larger than the Other, Ann. Math. Statist., 18, 50–60, https://doi.org/10.1214/aoms/1177730491, 1947.
Marshall, D. C.: Measurement of sap flow in conifers by heat transport, Plant Physiology, 33, 385–396, https://doi.org/10.1104/pp.33.6.385, 1958.
Marshall, J. D., Cuntz, M., Beyer, M., Dubbert, M., and Kuehnhammer, K.: Borehole Equilibration: Testing a New Method to Monitor the Isotopic Composition of Tree Xylem Water in situ, Frontiers in plant science, 11, 358, https://doi.org/10.3389/fpls.2020.00358, 2020.
Martín-Gómez, P., Serrano, L., and Ferrio, J. P.: Short-term dynamics of evaporative enrichment of xylem water in woody stems: implications for ecohydrology, Tree physiology, 37, 511–522, https://doi.org/10.1093/treephys/tpw115, 2017.
Marx, C., Tetzlaff, D., Hinkelmann, R., and Soulsby, C.: Seasonal variations in soil–plant interactions in contrasting urban green spaces: Insights from water stable isotopes, Journal of Hydrology, 612, 127998, https://doi.org/10.1016/j.jhydrol.2022.127998, 2022.
Meili, N., Manoli, G., Burlando, P., Carmeliet, J., Chow, W. T., Coutts, A. M., Roth, M., Velasco, E., Vivoni, E. R., and Fatichi, S.: Tree effects on urban microclimate: Diurnal, seasonal, and climatic temperature differences explained by separating radiation, evapotranspiration, and roughness effects, Urban Forestry & Urban Greening, 58, 126970, https://doi.org/10.1016/j.ufug.2020.126970, 2021.
Minick, K. J., Bahramian, J., Love, D., Tucker, L., Reinhardt, K., Johnson, D. M., and Emanuel, R. E.: Internal Water Movement and Residence Time Differ in Two Tree Species in a Temperate Deciduous Forest: Evidence From an In Situ D 2 O Isotope Tracer Study, Ecohydrology, 18, https://doi.org/10.1002/eco.70047, 2025.
Miralles, D. G., Gentine, P., Seneviratne, S. I., and Teuling, A. J.: Land-atmospheric feedbacks during droughts and heatwaves: state of the science and current challenges, Annals of the New York Academy of Sciences, 1436, 19–35, https://doi.org/10.1111/nyas.13912, 2019.
Nadezhdina, N., David, T. S., David, J. S., Ferreira, M. I., Dohnal, M., Tesaø, M., Gartner, K., Leitgeb, E., Nadezhdin, V., Cermak, J., Jimenez, M. S., and Morales, D.: Trees never rest: the multiple facets of hydraulic redistribution, Ecohydrology, 3, 431–444, https://doi.org/10.1002/eco.148, 2010.
O'Brien, J. J., Oberbauer, S. F., and Clark, D. B.: Whole tree xylem sap flow responses to multiple environmental variables in a wet tropical forest, Plant Cell & Environment, 27, 551–567, https://doi.org/10.1111/j.1365-3040.2003.01160.x, 2004.
Orlowski, N., Rinderer, M., Dubbert, M., Ceperley, N., Hrachowitz, M., Gessler, A., Rothfuss, Y., Sprenger, M., Heidbüchel, I., Kübert, A., Beyer, M., Zuecco, G., and McCarter, C.: Challenges in studying water fluxes within the soil-plant-atmosphere continuum: A tracer-based perspective on pathways to progress, The Science of the total environment, 881, 163510, https://doi.org/10.1016/j.scitotenv.2023.163510, 2023.
Penna, D., Hopp, L., Scandellari, F., Allen, S. T., Benettin, P., Beyer, M., Geris, J., Klaus, J., Marshall, J. D., Schwendenmann, L., Volkmann, T. H. M., von Freyberg, J., Amin, A., Ceperley, N., Engel, M., Frentress, J., Giambastiani, Y., McDonnell, J. J., Zuecco, G., Llorens, P., Siegwolf, R. T. W., Dawson, T. E., and Kirchner, J. W.: Ideas and perspectives: Tracing terrestrial ecosystem water fluxes using hydrogen and oxygen stable isotopes – challenges and opportunities from an interdisciplinary perspective, Biogeosciences, 15, 6399–6415, https://doi.org/10.5194/bg-15-6399-2018, 2018.
Petrova, I. Y., Miralles, D. G., Brient, F., Donat, M. G., Min, S.-K., Kim, Y.-H., and Bador, M.: Observation-constrained projections reveal longer-than-expected dry spells, Nature, 633, 594–600, https://doi.org/10.1038/s41586-024-07887-y, 2024.
Petruzzellis, F., Tordoni, E., Di Bonaventura, A., Tomasella, M., Natale, S., Panepinto, F., Bacaro, G., and Nardini, A.: Turgor loss point and vulnerability to xylem embolism predict species-specific risk of drought-induced decline of urban trees, Plant biology (Stuttgart, Germany), 24, 1198–1207, https://doi.org/10.1111/plb.13355, 2022.
Radolinski, J., Vremec, M., Wachter, H., Birk, S., Brüggemann, N., Herndl, M., Kahmen, A., Nelson, D. B., Kübert, A., Schaumberger, A., Stumpp, C., Tissink, M., Werner, C., and Bahn, M.: Drought in a warmer, CO2-rich climate restricts grassland water use and soil water mixing, Science (New York, N.Y.), 387, 290–296, https://doi.org/10.1126/science.ado0734, 2025.
Rahmani, F. and Fattahi, M. H.: A multifractal cross-correlation investigation into sensitivity and dependence of meteorological and hydrological droughts on precipitation and temperature, Nat. Hazards, 109, 2197–2219, https://doi.org/10.1007/s11069-021-04916-1, 2021.
R Core Team: _R: A Language and Environment for Statistical Computing_, R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org/ (last access: 15 August 2025), 2024.
Ring, A.: Key figure summarizing the sub-daily change in signatures of water stable isotopes in urban δxyl and δv (over grassland and under canopy). We compare summer drought (a) and autumn rewetting (b) in Berlin, Germany, 2022. Displayed amplitudes of δ²H [‰] were calculated from hourly medians from the 24-h cycle of each period, https://BioRender.com/s1f2bm9 (last access 12 August 2025), 2025.
Ring, A.-M., Tetzlaff, D., Dubbert, M., Dubbert, D., and Soulsby, C.: High-resolution in situ stable isotope measurements reveal contrasting atmospheric vapour dynamics above different urban vegetation, Hydrological Processes, 37, https://doi.org/10.1002/hyp.14989, 2023.
Ring, A.-M., Tetzlaff, D., Dubbert, M., Freymueller, J., and Soulsby, C.: Assessing the impact of drought on water cycling in urban trees via in-situ isotopic monitoring of plant xylem water, Journal of Hydrology, 633, 131020, https://doi.org/10.1016/j.jhydrol.2024.131020, 2024.
Roloff, A.: Bäume in der Stadt, Verlag Eugen Ulmer, ISBN 978-3-8001-7598-7, 2013.
Roloff, A., Korn, S., and Gillner, S.: The Climate-Species-Matrix to select tree species for urban habitats considering climate change, Urban Forestry & Urban Greening, 8, 295–308, https://doi.org/10.1016/j.ufug.2009.08.002, 2009.
Schweiger, A. H., Zimmermann, T., Poll, C., Marhan, S., Leyrer, V., and Berauer, B. J.: The need to decipher plant drought stress along the soil–plant–atmosphere continuum, Oikos, 2023, https://doi.org/10.1111/oik.10136, 2023.
Seeger, S. and Weiler, M.: Temporal dynamics of tree xylem water isotopes: in situ monitoring and modeling, Biogeosciences, 18, 4603–4627, https://doi.org/10.5194/bg-18-4603-2021, 2021.
SenMVKU: Percentage of public green spaces in Berlin, Grünflächeninformationssystem (GRIS), https://www.berlin.de/sen/uvk/_assets/natur-gruen/stadtgruen/daten-und-fakten/ausw_5.pdf (last access: 16 April 2025), 2023.
SenStadt (Berlin Senate Department for Urban Development, Building and Housing): Long-term Mean Precipitation Distribution 1991–2020, edited by: Haag, L., Berlin Environmental Atlas, https://www.berlin.de/umweltatlas/en/climate/precipitation-distribution/1991-2020/introduction/ (last access: 13 August 2025), 2023.
Shapiro, S. S. and Wilk, M. B.: An Analysis of Variance Test for Normality (Complete Samples), Biometrika, 52, 591, https://doi.org/10.2307/2333709, 1965.
Simonin, K. A., Roddy, A. B., Link, P., Apodaca, R., Tu, K. P., Hu, J., Dawson, T. E., and Barbour, M. M.: Isotopic composition of transpiration and rates of change in leaf water isotopologue storage in response to environmental variables, Plant Cell & Environment, 36, 2190–2206, https://doi.org/10.1111/pce.12129, 2013.
Smith, A., Tetzlaff, D., Landgraf, J., Dubbert, M., and Soulsby, C.: Modelling temporal variability of in-situ soil water and vegetation isotopes reveals ecohydrological couplings in a willow plot, Biogeosciences, 19, 2465–2485, https://doi.org/10.5194/bg-19-2465-2022, 2022.
Sobaga, A., Habets, F., Beaudoin, N., Léonard, J., and Decharme, B.: Decreasing trend of groundwater recharge with limited impact of intense precipitation: Evidence from long-term lysimeter data, Journal of Hydrology, 637, 131340, https://doi.org/10.1016/j.jhydrol.2024.131340, 2024.
Sohel, M. S. I., Herbohn, J., Nehemy, M. F., and McDonnell, J. J.: Differences between stem and branch xylem water isotope composition in four tropical tree species, Ecohydrology, 16, https://doi.org/10.1002/eco.2547, 2023a.
Sohel, M. S. I., Herbohn, J. L., Zhao, Y., and McDonnell, J. J.: Sap flux and stable isotopes of water show contrasting tree water uptake strategies in two co-occurring tropical rainforest tree species, Ecohydrology, 16, https://doi.org/10.1002/eco.2589, 2023b.
Steen-Larsen, H. C., Johnsen, S. J., Masson-Delmotte, V., Stenni, B., Risi, C., Sodemann, H., Balslev-Clausen, D., Blunier, T., Dahl-Jensen, D., Ellehøj, M. D., Falourd, S., Grindsted, A., Gkinis, V., Jouzel, J., Popp, T., Sheldon, S., Simonsen, S. B., Sjolte, J., Steffensen, J. P., Sperlich, P., Sveinbjörnsdóttir, A. E., Vinther, B. M., and White, J. W. C.: Continuous monitoring of summer surface water vapor isotopic composition above the Greenland Ice Sheet, Atmos. Chem. Phys., 13, 4815–4828, https://doi.org/10.5194/acp-13-4815-2013, 2013.
Steppe, K., Sterck, F., and Deslauriers, A.: Diel growth dynamics in tree stems: linking anatomy and ecophysiology, Trends in plant science, 20, 335–343, https://doi.org/10.1016/j.tplants.2015.03.015, 2015.
Stevenson, J. L., Birkel, C., Comte, J.-C., Tetzlaff, D., Marx, C., Neill, A., Maneta, M., Boll, J., and Soulsby, C.: Quantifying heterogeneity in ecohydrological partitioning in urban green spaces through the integration of empirical and modelling approaches, Environmental monitoring and assessment, 195, 468, https://doi.org/10.1007/s10661-023-11055-6, 2023.
Stevenson, J. L., Tetzlaff, D., Birkel, C., and Soulsby, C.: Contrasts in Ecohydrological Partitioning of Heterogeneous Urban Green Spaces in Energy-Limited Versus Water-Limited Hydroclimates, Hydrological Processes, 39, https://doi.org/10.1002/hyp.70077, 2025.
Tetzlaff, D., Buttle, J., Carey, S. K., McGuire, K., Laudon, H., and Soulsby, C.: Tracer-based assessment of flow paths, storage and runoff generation in northern catchments: a review, Hydrological Processes, 29, 3475–3490, https://doi.org/10.1002/hyp.10412, 2015.
Tetzlaff, D., Buttle, J., Carey, S. K., Kohn, M. J., Laudon, H., McNamara, J. P., Smith, A., Sprenger, M., and Soulsby, C.: Stable isotopes of water reveal differences in plant – soil water relationships across northern environments, Hydrological Processes, 35, https://doi.org/10.1002/hyp.14023, 2021.
Teuling, A. J., Seneviratne, S. I., Stöckli, R., Reichstein, M., Moors, E., Ciais, P., Luyssaert, S., van den Hurk, B., Ammann, C., Bernhofer, C., Dellwik, E., Gianelle, D., Gielen, B., Grünwald, T., Klumpp, K., Montagnani, L., Moureaux, C., Sottocornola, M., and Wohlfahrt, G.: Contrasting response of European forest and grassland energy exchange to heatwaves, Nature Geosci., 3, 722–727, https://doi.org/10.1038/ngeo950, 2010.
Teuling, A. J., van Loon, A. F., Seneviratne, S. I., Lehner, I., Aubinet, M., Heinesch, B., Bernhofer, C., Grünwald, T., Prasse, H., and Spank, U.: Evapotranspiration amplifies European summer drought, Geophysical Research Letters, 40, 2071–2075, https://doi.org/10.1002/grl.50495, 2013.
Tierney, K., Sobota, M., Snarski, J., Li, K., and Knighton, J.: Sub-Daily Variations in Tree Xylem Water Isotopic Compositions in a Temperate Northeastern US Forest, Hydrological Processes, 39, https://doi.org/10.1002/hyp.70137, 2025.
Umweltatlas Berlin: Flurabstand des Grundwassers 2020, https://www.berlin.de/umweltatlas/wasser/flurabstand/2020/karten/artikel.1322876.php (last access: 26 February 2025), 2020.
van Heerwaarden, C. C., Vilà-Guerau de Arellano, J., Moene, A. F., and Holtslag, A. A. M.: Interactions between dry-air entrainment, surface evaporation and convective boundary-layer development, Q. J. Roy. Meteor. Soc., 135, 1277–1291, https://doi.org/10.1002/qj.431, 2009.
van Loon, A. F.: Hydrological drought explained, WIREs Water, 2, 359–392, https://doi.org/10.1002/wat2.1085, 2015.
Vega-Grau, A. M., McDonnell, J., Schmidt, S., Annandale, M., and Herbohn, J.: Isotopic fractionation from deep roots to tall shoots: A forensic analysis of xylem water isotope composition in mature tropical savanna trees, The Science of the total environment, 795, 148675, https://doi.org/10.1016/j.scitotenv.2021.148675, 2021.
Vermunt, P. C., Steele-Dunne, S. C., Khabbazan, S., Judge, J., and van de Giesen, N. C.: Extrapolating continuous vegetation water content to understand sub-daily backscatter variations, Hydrol. Earth Syst. Sci., 26, 1223–1241, https://doi.org/10.5194/hess-26-1223-2022, 2022.
Volkmann, T. H. M. and Weiler, M.: Continual in situ monitoring of pore water stable isotopes in the subsurface, Hydrol. Earth Syst. Sci., 18, 1819–1833, https://doi.org/10.5194/hess-18-1819-2014, 2014.
Volkmann, T. H. M., Haberer, K., Gessler, A., and Weiler, M.: High-resolution isotope measurements resolve rapid ecohydrological dynamics at the soil-plant interface, The New phytologist, 210, 839–849, https://doi.org/10.1111/nph.13868, 2016a.
Volkmann, T. H. M., Kühnhammer, K., Herbstritt, B., Gessler, A., and Weiler, M.: A method for in situ monitoring of the isotope composition of tree xylem water using laser spectroscopy, Plant, cell & environment, 39, 2055–2063, https://doi.org/10.1111/pce.12725, 2016b.
Wang, H., Tetzlaff, D., Dick, J. J., and Soulsby, C.: Assessing the environmental controls on Scots pine transpiration and the implications for water partitioning in a boreal headwater catchment, Agricultural and Forest Meteorology, 240–241, 58–66, https://doi.org/10.1016/j.agrformet.2017.04.002, 2017.
Warix, S. R., Godsey, S. E., Flerchinger, G., Havens, S., Lohse, K. A., Bottenberg, H. C., Chu, X., Hale, R. L., and Seyfried, M.: Evapotranspiration and groundwater inputs control the timing of diel cycling of stream drying during low-flow periods, Front. Water, 5, https://doi.org/10.3389/frwa.2023.1279838, 2023.
Wassenaar, L. I., Hendry, M. J., Chostner, V. L., and Lis, G. P.: High resolution pore water delta2H and delta18O measurements by H2O(liquid)-H2O(vapor) equilibration laser spectroscopy, Environmental science & technology, 42, 9262–9267, https://doi.org/10.1021/es802065s, 2008.
Wei, Z., Yoshimura, K., Okazaki, A., Kim, W., Liu, Z., and Yokoi, M.: Partitioning of evapotranspiration using high-frequency water vapor isotopic measurement over a rice paddy field, Water Resources Research, 51, 3716–3729, https://doi.org/10.1002/2014WR016737, 2015.
Wilcoxon, F.: Individual Comparisons by Ranking Methods, Biometrics Bulletin, 1, 80, https://doi.org/10.2307/3001968, 1945.
Zannoni, D., Steen-Larsen, H. C., Sodemann, H., Thurnherr, I., Flamant, C., Chazette, P., Totems, J., Werner, M., and Raybaut, M.: Vertical and horizontal variability and representativeness of the water vapor isotope composition in the lower troposphere: insight from ultralight aircraft flights in southern France during summer 2021, Atmos. Chem. Phys., 25, 9471–9495, https://doi.org/10.5194/acp-25-9471-2025, 2025.
Zhang, Q., Manzoni, S., Katul, G., Porporato, A., and Yang, D.: The hysteretic evapotranspiration—Vapor pressure deficit relation, J. Geophys. Res.-Biogeo., 119, 125–140, https://doi.org/10.1002/2013JG002484, 2014.
Zhao, L., Wang, L., Cernusak, L. A., Liu, X., Xiao, H., Zhou, M., and Zhang, S.: Significant Difference in Hydrogen Isotope Composition Between Xylem and Tissue Water in Populus Euphratica, Plant, cell & environment, 39, 1848–1857, https://doi.org/10.1111/pce.12753, 2016.
Zhou, Y., Rosseau, G., Dao, V., and Wolfe, B. T.: Bark water vapor conductance varies among temperate forest tree species and is affected by flooding and stem bending, Tree physiology, 2024, tpae156, https://doi.org/10.1093/treephys/tpae156, 2024.